
Muscle Tissue: Anatomy, Physiology, and Clinical Applications
By the end of this exhaustively detailed guide, you will be able to:
- Understand the general characteristics and physiological properties of all muscle tissues.
- Differentiate the macroscopic and microscopic structural organization of skeletal, cardiac, and smooth muscle.
- Explain the precise molecular mechanisms of contraction across the three muscle types.
- Recognize the clinical and pathological implications of muscle tissue injury, regeneration, and malfunction.
1. Introduction to Muscle Tissue
Muscle tissue is a highly specialized, complex primary tissue type derived primarily from the embryonic mesoderm. It is composed of highly specialized, contractile cells that generate mechanical force to produce movement. The cells within all three types of muscle tissue are specialized for contraction (shortening), a dynamic process enabled by the ATP-driven interaction of specialized intracellular protein fibers (myofilaments).
This contraction enables the movement of the whole body (locomotion), the manipulation of the external environment, and the movement of substances through many internal hollow organs. In addition, muscle contraction is the primary mechanism for thermogenesis (producing heat energy) to maintain basal body temperature (e.g., shivering).
Cellular Division & Regeneration Constraint: An incredibly important characteristic of mature muscle cells is that they are generally terminally differentiated. They have entered the G0 phase of the cell cycle and have lost the ability to divide. Therefore, when muscle cells are heavily destroyed by trauma or ischemia (e.g., a heart attack), they often cannot be replaced by new functional muscle cells. Instead, the body repairs the defect with non-contractile fibrous connective tissue (scar tissue), a process known as fibrosis.
2. General Characteristics of All Muscle Tissue
Regardless of their location or specific type, all muscle tissues share a fundamental set of four key physiological properties that allow them to function effectively.
The ability to receive and respond to a stimulus. The stimulus is usually a chemical signal (like a neurotransmitter, e.g., Acetylcholine), a hormone, or a local change in pH. The response is the generation of an electrical impulse (action potential) that travels along the cell membrane.
The unique ability to shorten forcibly when adequately stimulated. This is what sets muscle apart from all other tissue types. Example: The biceps brachii contracting to lift a heavy weight.
The ability to be stretched or extended beyond their resting length without tearing. Example: The smooth muscle in the stomach wall stretching massively to accommodate a large holiday meal without bursting.
The ability of a muscle fiber to recoil and resume its original resting length after being stretched. This is governed by elastic proteins (like titin) within the cells. Example: The recoil of the heart muscle after it fills with blood.
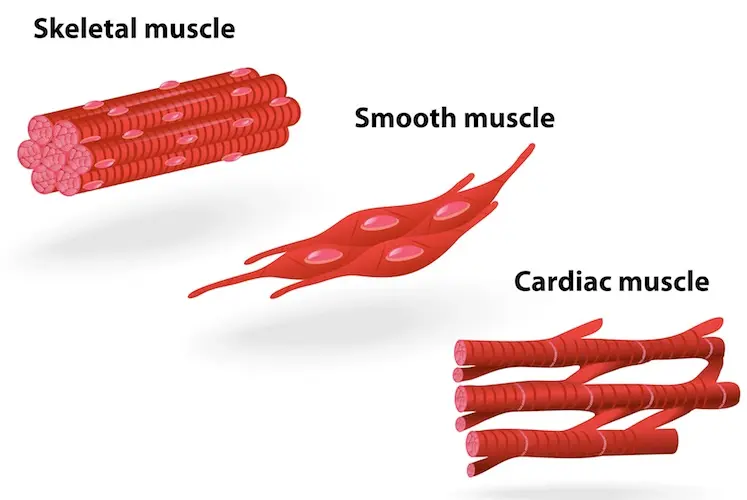
3. The Three Types of Muscle Tissue (Overview)
Muscle tissue is strictly classified into three types based on precise anatomical location, histological structure, and functional control characteristics. While the cells in smooth and cardiac muscle are referred to as muscle cells (myocytes), the exceptionally long, cylindrical cells of skeletal muscle are often called muscle fibers.
- 1. Skeletal Muscle: Striated (striped appearance), Voluntary (under conscious control), Multinucleate (long, cylindrical cells). Location: Primarily attached to the skeleton (bones) and skin, enabling gross body movement.
- 2. Cardiac Muscle: Striated (striped appearance), Involuntary (not under conscious control), Branched Cells (connected by specialized intercalated discs). Location: Found exclusively in the thick muscular wall of the heart (myocardium).
- 3. Smooth Muscle: Non-striated (smooth, uniform appearance), Involuntary (not under conscious control), Spindle-shaped Cells (featuring a single central nucleus). Location: Found in the walls of visceral hollow organs like the stomach, urinary bladder, uterus, and blood vessels.
4. Skeletal Muscle in Detail
Named for its anatomical location, skeletal muscle tissue is usually attached to bones via tendons, and sometimes directly to the skin (as in facial expression muscles). It enables voluntary movement of the head, trunk, and limbs. Its contractions are consciously controlled by the somatic nervous system.
Key Microscopic Characteristics:
- Striated: Under a light microscope, it appears distinctly striped or banded due to the highly organized, alternating arrangement of contractile proteins (actin and myosin).
- Voluntary: Contraction is consciously controlled by upper motor neurons originating in the cerebral cortex of the brain.
- Multi-nucleated Fibers: Because they are formed during embryonic development by the fusion of hundreds of smaller cells (myoblasts), a single mature skeletal muscle fiber is massive (up to 30 cm long) and contains hundreds of nuclei pushed to the extreme periphery of the cell just under the membrane.
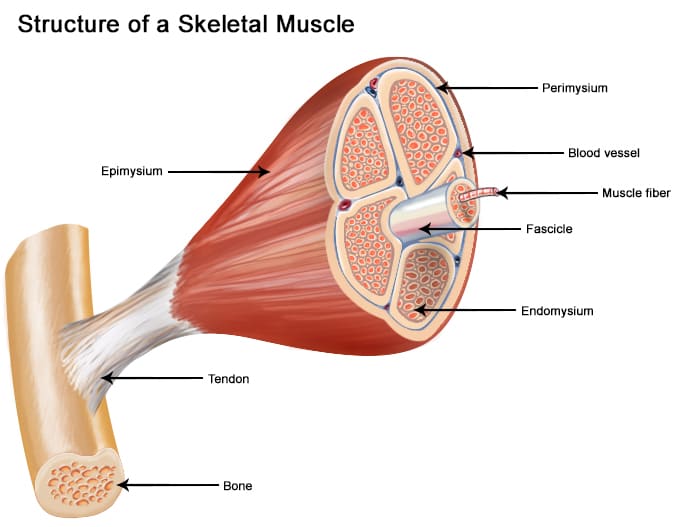
General Organization (Macroscopic to Microscopic)
A whole skeletal muscle is not just muscle cells; it is a complex organ containing muscle fibers, rich blood vessel networks, nerve fibers, and extensive connective tissue (CT) wrappings. These wrappings hold the fascicles together, provide passageways for blood vessels, and transmit the mechanical force of contraction to the bone.
- Entire Muscle (Organ level): The whole organ is surrounded by an outer sheath of dense irregular connective tissue called the Epimysium. This layer blends into the deep fascia surrounding other muscles.
- Muscle Fascicle: Inside the muscle, fibers are bundled together like a handful of dry spaghetti. Each bundle is called a fascicle. It is surrounded by a fibrous connective tissue layer called the Perimysium.
- Muscle Fiber (Cellular level): A single, exceptionally long muscle cell is individually wrapped by a delicate, fine layer of areolar connective tissue called the Endomysium.
Anatomical Note: All three of these "mysiums" are continuous with one another and converge at the ends of the muscle to form strong, rope-like tendons or broad, sheet-like aponeuroses, which anchor the muscle to the periosteum of the bone.
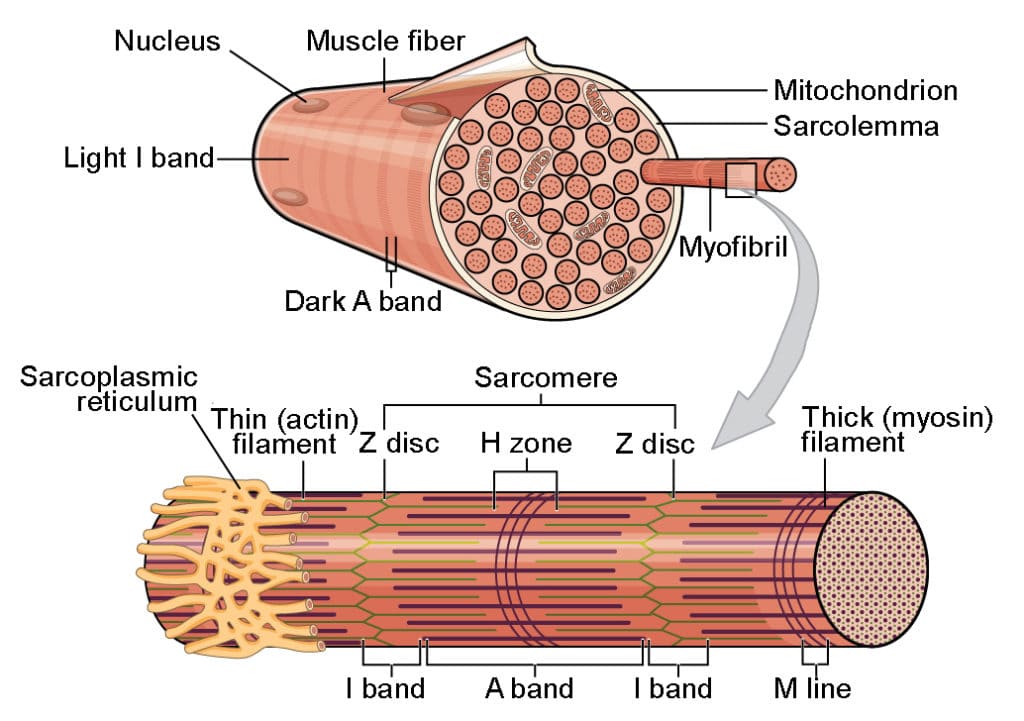
Microscopic Organization of a Muscle Fiber
A skeletal muscle fiber is a highly specialized cell optimized for rapid, explosive, and powerful contraction.
- Sarcolemma & Sarcoplasm: The Sarcolemma is the cell plasma membrane. It features specialized deep, tubular invaginations called T-tubules (Transverse tubules) that dive deep into the cell, ensuring electrical signals reach every part of the massive cell instantly. The Sarcoplasm is the specialized cytoplasm. It is uniquely rich in massive glycosomes (stored glycogen for instant glucose energy) and myoglobin (a red pigment that stores oxygen internally for the muscle's immediate use).
- Sarcoplasmic Reticulum (SR): A highly elaborate, specialized smooth endoplasmic reticulum that forms a web around each myofibril. Its primary, critical role is to store massive quantities of calcium ions (Ca²⁺) and release them on demand as the biological trigger for muscle contraction.
- Myofibrils and the Sarcomere: Inside the cell are thousands of rod-like contractile elements called Myofibrils (making up ~80% of the muscle fiber's total volume). Each myofibril is a long chain of repeating Sarcomeres, which are the fundamental, smallest functional contractile units of muscle.
The Sarcomere Banding Pattern (The Striations)
The striations you see under a microscope are actually the perfectly aligned boundaries of the sarcomeres:
- A-Band (Dark): Represents the full, exact length of the thick (myosin) filaments.
- I-Band (Light): Contains ONLY thin (actin) filaments.
- Z-Disc (Line): A dense zig-zag protein sheet that anchors the thin filaments and explicitly defines the exact boundaries of one single sarcomere (from Z-disc to Z-disc).
Myofilaments (The Contractile Proteins)
- Thick Filaments (Myosin): Composed of hundreds of myosin protein molecules bundled together. They feature flexible, globular "heads" that physically reach out to bind to actin and use ATP hydrolysis to generate pulling force.
- Thin Filaments (Actin): Composed of intertwined actin protein strands, which possess specific "active binding sites" for the myosin heads. The thin filaments also contain two critical regulatory proteins:
- Tropomyosin: A long, rope-like protein that acts as a physical shield. In a relaxed muscle, it physically blocks the myosin-binding sites on actin.
- Troponin: A molecular complex sitting on the tropomyosin. It acts as the "lock." It binds to Ca²⁺, which serves as the "key." When Ca²⁺ binds, troponin physically drags the tropomyosin rope out of the way, exposing the active sites and allowing contraction to begin.
The Sliding Filament Model of Contraction (Molecular Level)
This is the precise, step-by-step mechanism of how muscles shorten:
- Excitation: A motor nerve impulse arrives at the neuromuscular junction, releasing Acetylcholine (ACh). This sparks an electrical action potential that travels down the Sarcolemma and dives into the core of the cell via the T-tubules.
- Calcium Release: The electrical shock triggers the Sarcoplasmic Reticulum to instantly flood the sarcoplasm with Ca²⁺.
- Unshielding Actin: Ca²⁺ binds to troponin. Troponin changes shape and pulls tropomyosin away from actin's binding sites.
- Cross-Bridge Formation: Myosin heads (pre-energized by ATP) grab onto the newly exposed active sites on the actin.
- The Power Stroke: The myosin heads forcefully pivot and bend, physically dragging the thin filaments toward the center (M-line) of the sarcomere.
- Detachment & Reset: A new molecule of ATP must bind to the myosin head to force it to let go of the actin. The ATP is broken down (hydrolyzed), "re-cocking" the myosin head like a spring, ready for another cycle as long as Ca²⁺ and ATP are still present.
Result: The thick and thin filaments do not change length; they simply slide past one another. This sliding action shortens millions of sarcomeres simultaneously, causing the entire muscle organ to forcefully contract.
Rigor Mortis
When a person dies, their cells stop producing ATP. Shortly after death, calcium leaks out of the Sarcoplasmic Reticulum, exposing the actin binding sites. The myosin heads attach and pull, causing a muscular contraction. However, because the dead body has no ATP left to force the myosin heads to detach (Step 6 above), the cross-bridges become permanently locked. This results in the profound, total-body muscle stiffness known as Rigor Mortis, which only subsides days later when the muscle proteins literally begin to rot and decompose.
Satellite Cells and Muscle Repair
Since mature skeletal muscle fibers cannot divide, how do they grow when you lift weights, or repair when injured? The answer is Satellite Cells. These are quiescent (inactive) stem cells located tightly on the surface of mature muscle fibers, just beneath the endomysium. When a muscle fiber is traumatized (torn during a workout or injury), satellite cells are activated. They multiply and fuse with the existing muscle fibers to repair the damage and add new protein volume (hypertrophy). However, if the trauma is too massive, the satellite cells cannot keep up, and the gap is filled with fibrous scar tissue.
5. Cardiac Muscle Tissue
The muscle tissue located exclusively in the walls of the heart is cardiac muscle tissue. It consists of highly specialized branching cells that interconnect in a sophisticated netlike arrangement. The rhythmic, life-sustaining contractions of cardiac muscle are involuntary.
General Characteristics
- Location: Found exclusively in the Myocardium, the middle and thickest muscular layer of the heart wall.
- Function: Responsible for the forceful, rhythmic, tireless contractions that act as a biological pump, forcing blood throughout the vast cardiovascular network.
- Control: Strictly Involuntary. It possesses its own intrinsic, built-in electrical conduction system (autorhythmicity). The brain can speed the heart up or slow it down, but the heart generates its own beat independent of the brain.
- Appearance: It is striated, heavily resembling skeletal muscle, due to the identical organized arrangement of actin and myosin sarcomeres.
- Energy Needs: Has an extreme, continuous metabolic demand. It possesses abundant, gigantic mitochondria (taking up to 35% of the cell's volume, compared to 2% in skeletal muscle) and relies almost exclusively on aerobic (oxygen-based) respiration. The heart cannot survive on anaerobic energy; it will die rapidly without oxygen.
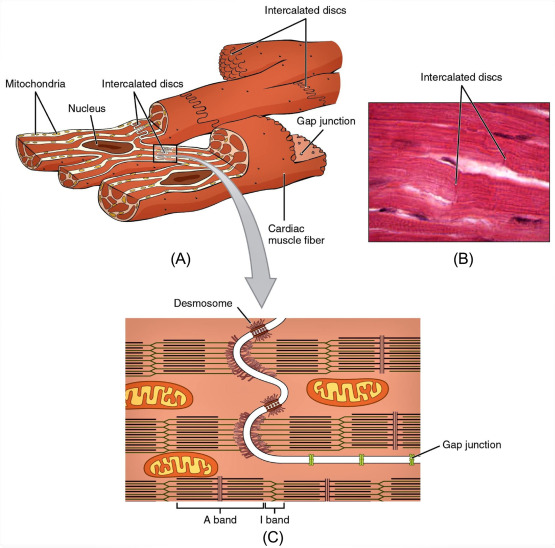
Microscopic Organization (The Cardiomyocyte)
Cardiac muscle cells, or cardiomyocytes, possess several unique morphological features tailored for collective cardiac function.
The Defining Feature: Intercalated Discs
These are complex, specialized microscopic junctions that uniquely connect adjacent cardiomyocytes end-to-end, appearing under a microscope as jagged, dark, wavy lines. They contain two vital structural components:
- Desmosomes: Act as biological rivets or heavy-duty molecular Velcro. They anchor the cells together, physically preventing them from tearing apart under the immense stress and pressure of the heart's forceful pumping action.
- Gap Junctions: Tiny, hollow protein channels that connect the cytoplasm of adjacent cells. They allow ions (and thus electrical action potentials) to pass directly and instantly from cell to cell. Because of gap junctions, the entire heart muscle acts and contracts as a single, coordinated unit—a phenomenon known as a functional syncytium.
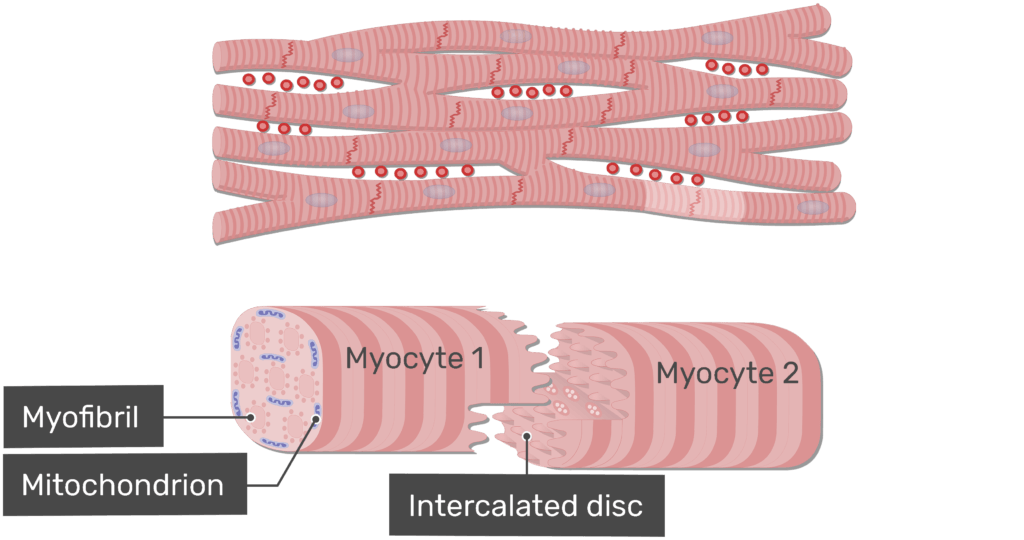
Other Microscopic Features:
- Cell Shape & Nuclei: Cells are significantly shorter, thicker, and highly branched (like the letter "Y"). Unlike skeletal muscle, most contain only a single, large, centrally located nucleus (occasionally two).
- Sarcoplasmic Reticulum (SR): Less extensive and less developed than in skeletal muscle. Cardiac muscle cannot store enough internal calcium, so it strictly relies on an influx of extracellular calcium to trigger contraction.
Mechanism of Contraction
Cardiac muscle contracts via the same sliding filament model (actin sliding over myosin), but with critical differences in how the contraction is initiated and regulated.
- Initiation (Autorhythmicity): Specialized pacemaker cells located in the Sinoatrial (SA) node spontaneously generate their own electrical impulses without any nervous system input. These impulses spread like a wave rapidly through the gap junctions to every cell in the heart.
- Calcium-Induced Calcium Release (CICR): When the electrical wave hits a cardiomyocyte, a small amount of extracellular Ca²⁺ enters the cell. This small drop of calcium triggers the Sarcoplasmic Reticulum to dump its massive stores of Ca²⁺ into the cell, which then unlocks troponin to allow contraction.
- The Long Refractory Period: Cardiac muscle has a massively prolonged "refractory period" (a time window where the cell simply refuses to be re-stimulated, lasting roughly 250 milliseconds).
Clinical Importance: This is a critical, life-saving safety feature. Because the refractory period is so long, cardiac muscle CANNOT undergo tetanic (sustained, locked) contractions. If the heart clamped down in tetanus, it would be unable to relax and refill with blood, causing instant death. The long refractory period guarantees the heart has time to relax and refill between every single beat.
Cardiac vs. Skeletal Muscle: Key Differences
| Feature | Skeletal Muscle | Cardiac Muscle |
|---|---|---|
| Control | Voluntary (Somatic nervous system) | Involuntary (Autorhythmic & Autonomic) |
| Cell Shape | Very long, cylindrical, unbranched | Shorter, highly branched |
| Nuclei | Many, pushed to the periphery | One or two, centrally located |
| Intercalated Discs | Absent | Present (Desmosomes & Gap Junctions) |
| Ca²⁺ Source | Almost entirely from internal SR stores | SR + critical influx of Extracellular Ca²⁺ |
| Refractory Period | Short (can go into sustained tetanus) | Extremely Long (tetanus is impossible) |
| Mitochondria | ~2% of total cell volume | 25-35% of total cell volume |
6. Smooth Muscle Tissue
Smooth muscle is morphologically and functionally very distinct from both skeletal and cardiac muscle. It is specialized for exceptionally slow, sustained, highly energy-efficient involuntary contractions. It is intimately involved in maintaining the internal physical environment (homeostasis) and is primarily found in the walls of hollow internal visceral organs.
General Characteristics
- Location: Forms the muscular walls of hollow organs like the digestive tract (esophagus, stomach, intestines), urinary bladder, reproductive tract (uterus), blood vessels (arteries and veins), and the respiratory airways (bronchioles).
- Function: Propulsion of substances via wave-like contractions (peristalsis), regulation of blood flow and blood pressure (vasoconstriction and vasodilation), and forceful expulsion of organ contents (urination or childbirth).
- Control: Strictly Involuntary. It is heavily regulated by the Autonomic Nervous System (ANS), circulating hormones (like oxytocin or adrenaline), and local chemical changes (like low oxygen or high acidity).
- Appearance: Non-striated. It appears smooth and uniform under a microscope because its contractile proteins are not arranged into perfectly organized, striped sarcomeres.
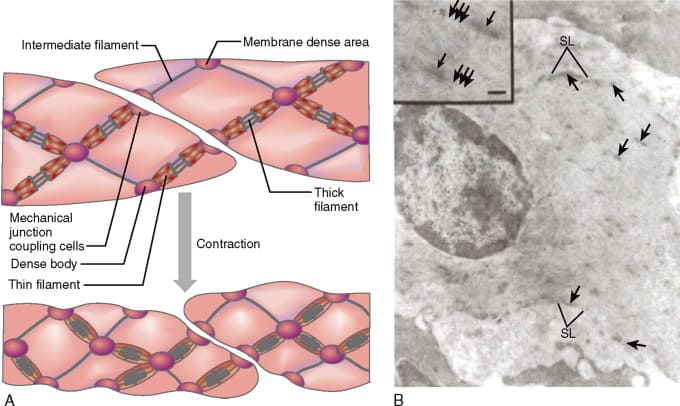
Microscopic Organization (The Leiomyocyte)
Smooth muscle cells (leiomyocytes) are relatively simple in their morphology but possess an incredibly sophisticated and unique contractile mechanism.
- Spindle-shaped Cells: The cells are elongated, widest in the middle, and neatly taper off to sharp, pointed ends. Each cell contains a single, cigar-shaped, centrally located nucleus.
- No Sarcomeres: Actin and myosin filaments are definitely present, but they are arranged diagonally in a criss-crossing, lattice-like network. They are anchored to the cell membrane by structures called Dense Bodies (which are the smooth muscle equivalent of Z-discs). When the cell contracts, it physically twists and corkscrews, wringing out like a wet towel.
- Calcium Source: The sarcoplasmic reticulum (SR) is extremely poorly developed. Therefore, the vast majority of the Ca²⁺ required for contraction must flood in from the Extracellular Fluid (ECF) via calcium channels on the cell membrane.
- No Troponin: Smooth muscle lacks the troponin complex entirely. Instead, to initiate contraction, the incoming Ca²⁺ binds to a completely different regulatory protein called Calmodulin.
Because there is no troponin, smooth muscle contraction works via a unique biochemical cascade:
- Calcium enters the cell and binds to Calmodulin.
- The Calcium-Calmodulin complex activates a special enzyme called Myosin Light Chain Kinase (MLCK).
- Activated MLCK takes a phosphate from ATP and attaches it directly to the myosin head.
- Only when phosphorylated can the myosin head bind to actin and perform the power stroke.
Efficiency Note (The Latch-Bridge Mechanism): Smooth muscle is incredibly energy-efficient. Once the myosin attaches to actin, it can enter a "latch state," maintaining severe tension for hours while using barely any ATP. This allows blood vessels to maintain constant blood pressure all day without exhausting themselves.
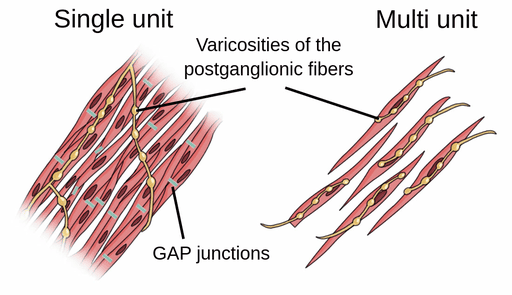
Types of Smooth Muscle
Smooth muscle is broadly categorized into two completely different types based on its neural wiring and functional characteristics:
The overwhelmingly most common type. The cells are electrically coupled tightly together by millions of gap junctions. Because they share electrical signals instantly, the entire sheet of muscle contracts rhythmically and simultaneously as a single unit (another example of a functional syncytium). Location & Example: Found in the walls of the stomach and intestines to coordinate the massive sweeping waves of peristalsis.
Consists of individual, structurally independent cells with few or absolutely no gap junctions. Each individual cell has its own dedicated nerve ending. This setup allows for fine, incredibly precise, graded control, similar to skeletal muscle. Location & Example: Found in the large airways of the lungs, the walls of large arteries, the tiny piloerector muscles that cause goosebumps, and the iris of the eye to precisely control pupil dilation.
Smooth vs. Striated Muscle: Key Differences
| Feature | Striated Muscle (Skeletal & Cardiac) | Smooth Muscle |
|---|---|---|
| Striations | Yes (due to perfectly aligned sarcomeres) | No (no organized sarcomeres) |
| Control | Voluntary (Skeletal) / Involuntary (Cardiac) | Strictly Involuntary |
| Cell Shape | Long, cylindrical or branched | Spindle-shaped (fusiform) |
| Calcium Binding | Troponin complex binds Ca²⁺ | Calmodulin binds Ca²⁺ |
| Ca²⁺ Source | Primarily SR (Skeletal), SR + ECF (Cardiac) | Primarily Extracellular Fluid (ECF) |
| Contraction Speed | Fast, rapid, and explosive | Very slow, sustained, and prolonged |
| Fatigue Resistance | Moderate (Skeletal), High (Cardiac) | Extremely High (Latch-bridge mechanism) |
References
- Hall, J. E., & Hall, M. E. (2020). Guyton and Hall Textbook of Medical Physiology (14th ed.). Elsevier.
- Marieb, E. N., & Hoehn, K. (2018). Human Anatomy & Physiology (11th ed.). Pearson.
- Tortora, G. J., & Derrickson, B. (2017). Principles of Anatomy and Physiology (15th ed.). Wiley.
- Mescher, A. L. (2018). Junqueira's Basic Histology: Text and Atlas (15th ed.). McGraw-Hill Education.
- Costanzo, L. S. (2017). Physiology (6th ed.). Elsevier.