
Nerve and Muscle Physiology
By the conclusion of this exhaustive master guide, you will be deeply conversant with:
- The fundamental properties of excitable tissues and how voltage gradients dictate human physiology.
- The precise structural and functional anatomy of the motor neuron.
- The step-by-step biochemical and electrical cascade of synaptic transmission and the generation of an action potential.
- The microscopic anatomy of skeletal muscle, including the structural hierarchy of the sarcomere.
- The flawless sequence of Excitation-Contraction Coupling, from the Neuromuscular Junction (NMJ) down to the exact molecular movements of the Sliding Filament Theory.
I. Introduction to Excitable Tissues
Nerve and muscle physiology is a highly specialized branch of medical physiology that specifically studies the function, architecture, and electrochemical mechanisms of nervous tissue (nerves) and muscle tissue (muscles). Collectively, nerves and muscles are uniquely classified as "excitable tissues."
This classification means they possess the extraordinary ability to undergo rapid, transient changes in their resting membrane potential. They explore how these tissues generate and transmit electrical signals (like action potentials) and how these sheer electrical impulses are seamlessly converted into specific, tangible cellular functions.
- For Nerves: It covers how neurons (individual nerve cells) generate electrical impulses, communicate with each other across microscopic gaps (synaptic transmission), process immense amounts of sensory information, and transmit command signals throughout the body to control various systemic functions—ranging from conscious thought and pain sensation to voluntary movement and involuntary organ regulation.
- For Muscles: It focuses intensely on how muscle cells (muscle fibers) respond to the electrical signals delivered by nerves, leading to mechanical contraction (physical shortening) and the generation of kinetic force. This includes the intricate molecular mechanisms of contraction, the metabolic regulation of muscle force, and the different types of muscle tissue and their distinct, highly specialized functional characteristics.
Nervous System Excitability
Nervous system excitability is defined as the inherent ability of nerve cells (neurons) to respond to an external or internal stimulus by generating and propagating an action potential—a massive, self-propagating electrical impulse that travels down the length of the cell.
This property is the absolute fundamental basis of the nervous system's function. It depends entirely on the neuron membrane's selective permeability, the precise opening and closing of voltage-gated ion channels, and the relentless work of active ion pumps. A sudden, sufficient change in membrane potential leads directly to this firing event, which is essential for transmitting information throughout the entire body. The physiology of the nervous system involves its main divisions—the Central Nervous System (CNS) and Peripheral Nervous System (PNS)—which utilize neurons and electrochemical signals to sense stimuli, integrate complex data, and produce highly coordinated, life-sustaining responses.
II. Overall Structure & Function of a Motor Neuron (The Command Pathway)
A motor neuron is a highly specialized, efferent nerve cell that transmits electrical command signals strictly away from the central nervous system (brain and spinal cord) to effector targets like muscles or glands, thereby initiating physical movement or chemical secretion.
The "Final Common Pathway"
Motor neurons are universally referred to in physiology as the final common pathway. This term emphasizes a profound physiological principle: all the impossibly complex neural computations happening in higher brain centers (e.g., motor planning in the cerebral cortex, coordination in the basal ganglia, and balance checking in the cerebellum) must ultimately converge onto these lower motor neurons.
It is only through the firing of a lower motor neuron that a skeletal muscle can be activated and a movement can physically occur. Regardless of whether a movement is voluntary (throwing a ball) or reflexive (pulling your hand away from a hot stove), the command signal ultimately travels down a lower motor neuron to its target muscle fibers. This makes the motor neuron a critical biological bottleneck and the ultimate determinant of muscle activity.
Clinical Example: Amyotrophic Lateral Sclerosis (ALS) is a devastating disease that specifically targets and destroys these motor neurons. Because they are the final common pathway, their destruction leads to complete, irreversible muscle paralysis, even though the patient's higher brain functions remain perfectly intact.

1. Motor Neuron Anatomy: Key Structural Components
The metabolic and genetic center of the neuron. It contains the nucleus, extensive rough endoplasmic reticulum (Nissl bodies), and other organelles. It synthesizes vital neurotransmitters and structural proteins, and it receives synaptic inputs directly from thousands of other neurons.
Branching, tree-like extensions protruding from the soma. These are the primary receptive (input) regions of the cell. They are studded with ligand-gated ion channels that receive chemical signals (neurotransmitters) from adjacent cells and convert them into graded electrical potentials (EPSPs and IPSPs).
A specialized, cone-shaped anatomical region where the axon originates from the soma. This is the critical "trigger zone". It possesses the highest density of voltage-gated Na⁺ channels in the entire cell. It acts as an organic calculator, mathematically integrating all incoming graded potentials; if the sum reaches the critical threshold, an action potential is violently generated here.
A single, microscopic, tube-like projection that transmits the action potential (the output signal) away from the cell body toward the target. Some axons (like those reaching down to your toes) can exceed a full meter in length!
A thick, fatty, insulating layer wrapped around many axons. It is formed by Schwann cells in the PNS and Oligodendrocytes in the CNS. It is absolutely crucial for dramatically increasing the speed of action potential conduction and preventing electrical signal leakage.
Microscopic, unmyelinated gaps evenly spaced along the myelin sheath. These exposed patches contain a massive concentration of voltage-gated Na⁺ and K⁺ channels. The action potential is chemically regenerated at these nodes, allowing the signal to "jump" lightning-fast from node to node in a process called saltatory conduction.
The highly branched, club-like ends of the axon that form physical synapses with other cells. They contain thousands of synaptic vesicles packed with neurotransmitters and are specialized for converting the electrical action potential back into a chemical signal.
2. Functional Zones: Relating Structure to Role
To understand the flow of neurological information, we map these anatomical structures into four distinct functional zones:
- Input Zone (Dendrites & Cell Body): Receives and chemically integrates incoming signals, translating them into graded potentials.
- Integration Zone (Axon Hillock): The decision-making center. It sums all graded potentials. If the net depolarization is strong enough to reach threshold, it fires the weapon (triggers an action potential).
- Conduction Zone (Axon): Propagates the "all-or-nothing" action potential without any loss of signal strength over immense distances, heavily facilitated by the myelin sheath's saltatory conduction.
- Output Zone (Axon Terminals): Converts the traveling electrical action potential into a chemical signal by dumping neurotransmitters onto the next cell.
III. Synaptic Transmission: The Communication Bridge
Synaptic transmission is the fundamental, microscopic process by which one neuron (the presynaptic neuron) communicates with another neuron (the postsynaptic neuron) or an effector cell (like a muscle). Most synapses in the human nervous system are chemical synapses, meaning they utilize chemical messengers known as neurotransmitters to physically bridge the gap between cells where electricity cannot cross.

Anatomy of a Chemical Synapse
- Presynaptic Terminal (Axon Terminal): The specialized emitting end of the incoming axon. It is densely packed with synaptic vesicles (bubbles full of neurotransmitters), abundant mitochondria to fuel the constant energy demand, and crucial voltage-gated Ca²⁺ channels.
- Synaptic Cleft: The microscopic, fluid-filled extracellular void (typically only 20-50 nanometers wide) that physically separates the sending and receiving membranes.
- Postsynaptic Membrane: The highly specialized receiving region of the target cell's membrane, heavily studded with specific neurotransmitter receptors.
The Step-by-Step Mechanism of Neurotransmission
1. Presynaptic Events: Neurotransmitter Release
This phase is the miracle of converting an electrical spark into a chemical flood:
- Action Potential Arrives: An action potential successfully propagates down the axon and violently depolarizes the presynaptic terminal membrane.
- Depolarization Opens Voltage-Gated Ca²⁺ Channels: The sudden shift in positive electrical charge forces specialized calcium channels in the terminal membrane to snap open.
- Ca²⁺ Influx: Driven by a steep electrochemical gradient (calcium is highly concentrated outside the cell), Ca²⁺ ions rapidly rush into the presynaptic terminal. This influx is the absolute, non-negotiable trigger for neurotransmitter release.
- Ca²⁺ Triggers Vesicle Fusion: The sudden spike in intracellular Ca²⁺ activates specific docking proteins (SNARE proteins). These proteins act like winches, pulling the synaptic vesicles down and forcing them to fuse with the presynaptic cell membrane.
- Neurotransmitter Release (Exocytosis): As the vesicles violently fuse with the membrane, they burst open, expelling their payload of neurotransmitters directly into the synaptic cleft.
Botulism & Tetanus Toxins
The deadly Botulinum toxin (used cosmetically as Botox) and Tetanus toxin work by destroying the SNARE proteins mentioned in Step 4. Without SNARE proteins, synaptic vesicles cannot fuse with the membrane. The nerve fires, calcium rushes in, but the neurotransmitters are permanently trapped inside the terminal. This completely severs communication to the muscle, resulting in flaccid paralysis (in Botulism) or rigid, spastic paralysis (in Tetanus).
2. Postsynaptic Events: Receptor Binding & Ion Channel Opening
Once dumped into the cleft, neurotransmitters diffuse rapidly across the microscopic gap and bind reversibly to their specific, lock-and-key receptors on the postsynaptic membrane.
- Ligand-Gated Ion Channels (Ionotropic): The receptor itself is a physical ion channel. Binding of the neurotransmitter acts as a key that instantly pops the channel open, allowing an immediate rush of ions and a rapid change in the postsynaptic membrane potential. This generates:
- Excitatory Postsynaptic Potential (EPSP): A localized depolarization (e.g., via Na⁺ influx), making the inside of the neuron more positive and therefore more likely to fire.
- Inhibitory Postsynaptic Potential (IPSP): A localized hyperpolarization (e.g., via Cl⁻ influx or K⁺ efflux), making the inside of the neuron more negative and effectively "shutting it down," making it less likely to fire.
- G-Protein Coupled Receptors (Metabotropic): The receptor activates an intracellular middle-man called a G-protein. This initiates a slower, highly complex, but much more widespread and long-lasting signaling cascade within the cell. This can lead to the production of "second messengers" (e.g., cAMP) that can physically alter the cell's DNA gene expression or modulate nearby ion channels from the inside.
3. Neurotransmitter Inactivation/Removal: Terminating the Signal
To ensure precise, crisp, and discrete signaling, the chemical message must be swiftly deleted from the cleft the moment the signal is delivered. If left unchecked, the muscle or nerve would seize up in a state of permanent, toxic overstimulation. This termination happens through three mechanisms:
- Enzymatic Degradation: Specific assassin enzymes localized in the synaptic cleft chemically chop up and destroy the neurotransmitter. Example: Acetylcholinesterase (AChE) brutally breaks down acetylcholine into harmless choline and acetate in milliseconds.
- Reuptake: Specialized, ATP-driven transporter proteins on the presynaptic terminal (or nearby glial cells) actively act like biological vacuum cleaners, pumping the intact neurotransmitter back into the sending cell to be recycled for the next firing. Example: This is the primary mechanism for monoamines like serotonin and dopamine.
Clinical Application: Selective Serotonin Reuptake Inhibitors (SSRIs) like Prozac block these vacuum pumps, intentionally leaving serotonin in the cleft longer to treat severe depression. - Diffusion: Some neurotransmitters simply float and diffuse away from the synaptic cleft into the surrounding extracellular fluid, where their concentration drops to ineffective levels.

IV. Generation and Conduction of the Action Potential
The motor neuron is constantly bombarded with thousands of chemical signals (both excitatory and inhibitory) from thousands of other neurons every second. The neuron must mathematically integrate these signals to decide whether to fire a massive, "all-or-nothing" action potential.
1. Integration: Spatial and Temporal Summation
A single tiny EPSP is overwhelmingly too weak to push a neuron to threshold. Therefore, the Axon Hillock adds them all up through summation:
- Spatial Summation: Multiple EPSPs or IPSPs arriving at completely different physical locations on the dendrites at the exact same time add together. (Analogy: Ten different people pushing a heavy car at the same time).
- Temporal Summation: Rapid, successive, machine-gun-like EPSPs from a single presynaptic neuron add up over time before the previous one has a chance to fade away. (Analogy: One person rhythmically pushing a swing over and over to build momentum).
If the absolute algebraic sum of all incoming excitatory (EPSPs) and inhibitory (IPSPs) signals reaches the critical Threshold Potential (typically around -55 mV), the Axon Hillock opens its gates and an action potential is irrevocably generated.
2. Phases of the Action Potential
Once generated, the action potential propagates down the axon without losing a fraction of its strength. It follows a highly stereotyped, predictable voltage curve:
| Phase | Voltage Level | Cellular Event (Ion Channel Status) |
|---|---|---|
| 1. Resting State | -70 mV | The membrane is highly polarized. All voltage-gated Na⁺ and K⁺ channels are tightly closed. The Resting Membrane Potential (RMP) is maintained primarily by K⁺ leak channels (potassium constantly trickling out) and the relentless, energy-burning Na⁺/K⁺-ATPase pump. |
| 2. Depolarization to Threshold | -70 mV to -55 mV | The summed EPSPs cause a localized, sluggish depolarization. If the internal charge reaches the -55 mV threshold, it triggers a catastrophic, positive feedback loop. |
| 3. Rising Phase (Depolarization) | -55 mV shooting up to +30 mV | At threshold, thousands of voltage-gated Na⁺ channels violently snap open. A massive, sudden influx of Na⁺ rushes into the cell, erasing the negative charge and making the inside of the membrane highly positive. |
| 4. Falling Phase (Repolarization) | +30 mV dropping back toward -70 mV | At the exact peak (+30 mV), an inactivation gate swings shut on the Na⁺ channels, instantly stopping the Na⁺ influx. Simultaneously, the sluggish voltage-gated K⁺ channels finally fully open. A massive efflux of K⁺ rushes out of the cell, rapidly repolarizing the membrane back to negative. |
| 5. Undershoot (Hyperpolarization) | Dips below -70 mV (e.g., -80 mV) | The voltage-gated K⁺ channels are slow to close, causing an excessive, prolonged efflux of K⁺. The membrane becomes briefly more negative than its resting state. During this time, the neuron enters its Relative Refractory Period, where only an abnormally massive stimulus can trigger another firing. (Note: The Absolute Refractory Period occurs during the rising and falling phases, where it is physically impossible for the cell to fire again, ensuring the signal only travels in one forward direction). |
| 6. Restoration | Returns to -70 mV | All voltage-gated channels finally slam shut. The Na⁺/K⁺-ATPase pump works continuously in the background, burning ATP to shovel Na⁺ back out and K⁺ back in, fully restoring the long-term chemical gradients. |
Local Anesthetics (Lidocaine)
When a dentist injects Lidocaine into your gums, the drug physically enters the nerve cell and plugs up the Voltage-Gated Na⁺ Channels from the inside. If Na⁺ cannot rush in during the Rising Phase, the action potential completely fails to generate. The pain signal is stopped dead in its tracks, and your brain never feels the drill.
V. Muscle Physiology: The Architecture of Contraction
While nerves specialize in communication, muscle tissue is purely specialized for mechanical contraction, generating kinetic force and bodily movement. We focus here on Skeletal Muscle.
A. Skeletal Muscle Micro-Structure
- Muscle Fiber (Cell): A single, highly elongated, cylindrical, multinucleated cell spanning the length of the muscle.
- Sarcolemma: The specialized, electrically excitable plasma membrane of a muscle fiber. It features deep, tube-like invaginations that dive into the center of the cell called T-tubules.
- Sarcoplasm: The unique cytoplasm of a muscle fiber. It is heavily packed with mitochondria (for ATP), glycogen (sugar storage), myoglobin (oxygen storage), and thousands of contractile rods called myofibrils.
- Myofibrils: Long, rod-like contractile organelles running parallel to the fiber, entirely composed of repeating microscopic units called sarcomeres.
- Sarcoplasmic Reticulum (SR): A highly specialized, web-like smooth endoplasmic reticulum that wraps around each and every myofibril like a sleeve. Its sole purpose is storing, releasing, and recapturing massive amounts of Ca²⁺ ions.
- The Triad: A critical anatomical junction consisting of one central T-tubule physically flanked on both sides by two enlarged sacs of the SR (terminal cisternae). This ensures the electrical signal diving down the T-tubule instantly triggers the adjacent SR.

B. The Sarcomere: The Ultimate Contractile Unit
The sarcomere is the fundamental, repeating contractile unit of a myofibril, extending from one Z-disc to the next Z-disc. Its highly organized geometry gives skeletal muscle its striated (striped) appearance.
The Filaments (The Machinery):
- Thick Filaments (Myosin): Composed entirely of the protein myosin. Each myosin molecule looks like a golf club, with a twisted tail and two globular, pivoting heads. The heads are the true engines; they contain an actin-binding site and an ATP-binding site (which functions as an ATPase enzyme to burn fuel).
- Thin Filaments (Actin Complex): Composed primarily of a twisted double-strand of actin pearls. Critically, the thin filament also houses two highly sensitive regulatory proteins that act as a lock-and-key system:
- Tropomyosin: A long, thread-like, rod-shaped protein that spirals around the actin. In a relaxed muscle, it physically covers and blocks the myosin-binding sites on the actin, preventing contraction.
- Troponin: A specialized complex of three proteins pinned to tropomyosin. The Troponin C (TnC) subunit acts as the ultimate lock; it is the specific component that strongly binds incoming Ca²⁺ ions, shifting the tropomyosin out of the way and initiating the violent contraction sequence.
The Bands and Zones (The Geography):
- A Band: The entire, dark length of the thick (myosin) filament. Crucial rule: Its physical length remains absolutely constant and never changes during contraction.
- I Band: The light region containing only thin (actin) filaments. It drastically shortens during contraction as the filaments slide inward.
- H Zone: The lighter, central region of the A band containing only thick filaments (no actin overlap). It completely disappears/shortens during maximum contraction.
- M Line: A dark, structural anchoring line running straight down the dead center of the H zone, holding the thick filaments in strict alignment.
- Z Disc (Z Line): The jagged, zig-zag boundary line that perfectly defines the absolute ends of a single sarcomere and anchors the thin filaments in place.

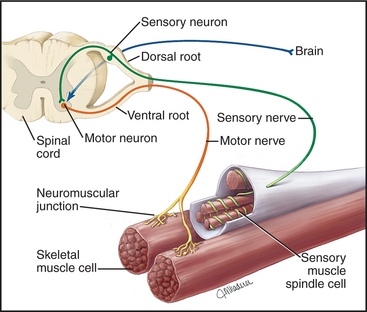
VI. The Neuromuscular Junction (NMJ): The Nerve-Muscle Interface
The neuromuscular junction (NMJ) is arguably the most important chemical synapse in the human body. It is the highly specialized, mandatory interface where a motor neuron's axon terminal docks with a skeletal muscle fiber.
Anatomy of the NMJ:
- Presynaptic Terminal: The swollen end of the motor neuron's axon, packed tight with thousands of synaptic vesicles heavily loaded with the neurotransmitter Acetylcholine (ACh).
- Synaptic Cleft: The microscopic physical gap between the nerve and the muscle. It is flooded with a highly active enzyme called Acetylcholinesterase (AChE).
- Motor End Plate: A highly specialized, crater-like region of the muscle's sarcolemma positioned directly under the nerve terminal. It features deep, accordion-like junctional folds densely packed with millions of specialized Nicotinic Acetylcholine Receptors (nAChRs).
End-Plate Potential (EPP): The Muscle's First Electrical Response
This is the explosive, step-by-step translation of a nerve thought into a muscle shock:
- ACh Release: An action potential traveling down the motor neuron opens terminal Ca²⁺ channels, triggering the massive exocytosis of ACh into the synaptic cleft.
- ACh Binding: ACh diffuses across the tiny cleft and instantly binds to the nAChR receptors waiting on the motor end plate.
- Channel Opening: The binding of exactly two ACh molecules to a receptor forces the central ion channel to pop open.
- Ion Movement: Na⁺ ions rapidly rush into the muscle fiber, while a smaller, slower stream of K⁺ ions moves out. The absolute net effect is a massive influx of positive charge into the muscle.
- Depolarization (EPP): This sudden net influx of positive Na⁺ ions causes a rapid, incredibly large, localized depolarization of the motor end plate, scientifically known as the End-Plate Potential (EPP). Important physiological note: Unlike the weak EPSPs in the brain, a single healthy EPP is immensely powerful and is always large enough to instantly trigger a full, unstoppable action potential in the adjacent sarcolemma (this is known as the safety factor).
- ACh Inactivation: In mere milliseconds, the ACh is violently ripped off the receptor and degraded by the enzyme acetylcholinesterase (AChE) suspended in the cleft, terminating the excitatory signal instantly and allowing the muscle fiber to repolarize for the next breath or step.
Myasthenia Gravis vs. Nerve Gas
- Myasthenia Gravis: An autoimmune disease where the body mistakenly produces antibodies that attack and destroy the nAChRs on the motor end plate. Even though the nerve releases plenty of ACh, there are no receptors left to catch it. The EPP fails to reach threshold, resulting in profound, progressive muscle weakness (drooping eyelids, inability to swallow).
- Sarin Nerve Gas & Organophosphates: These toxic chemicals permanently destroy the enzyme AChE in the synaptic cleft. ACh is released but is never destroyed. It violently overstimulates the receptors continuously, causing the muscles to undergo fatal, tetanic, spastic paralysis (the diaphragm locks up and the victim suffocates).
VII. Excitation-Contraction Coupling
This is the awe-inspiring physiological bridge by which an invisible electrical signal (the muscle action potential) is instantly converted into a violent mechanical event (muscle contraction).
- Muscle AP Propagation: The newly generated action potential travels like a wave of fire across the surface of the sarcolemma and rapidly dives deep down into the core of the cell via the T-tubules.
- DHPR Activation: As the electrical shock travels down the T-tubule, it hits and alters the physical shape of specialized voltage-sensitive sensor proteins in the membrane called Dihydropyridine Receptors (DHPRs).
- Mechanical Linkage to RyRs: In skeletal muscle, these DHPR sensors are physically, mechanically linked like a locked door handle to Ryanodine Receptors (RyRs). The RyRs are giant "plug" channels embedded directly in the membrane of the adjacent Sarcoplasmic Reticulum (SR).
- RyR Opening and Ca²⁺ Release: The voltage change forces the DHPR to literally pull the RyR plug open mechanically. This unplugs the SR, allowing billions of stored, highly pressurized Ca²⁺ ions to violently flood out of the SR and into the surrounding sarcoplasm (cytoplasm).
- Increase in Intracellular Ca²⁺: This sudden, massive spike in sarcoplasmic Ca²⁺ concentration is the absolute, immediate, non-negotiable chemical trigger for muscle contraction.

VIII. The Mechanism of Muscle Contraction: The "Sliding Filament Theory"
The Sliding Filament Theory universally proposes that muscle shortening (contraction) occurs strictly by the thick (myosin) and thin (actin) filaments sliding past one another, dragging the Z-discs closer together. The filaments themselves never actually shrink or change length; they merely increase their overlap.
1. Role of Ca²⁺: Unlocking the Binding Sites
Before the engines can engage, the track must be cleared. The Ca²⁺ ions released from the SR flood the sarcomere and bind immediately to the Troponin C subunit perched on the thin actin filaments. This binding causes a profound shape change in the entire troponin complex, which in turn violently tugs on the long, thread-like tropomyosin molecule. The movement of tropomyosin physically drags it away from the myosin-binding sites on the actin beads, exposing them completely to the waiting myosin heads.

2. The Cross-Bridge Cycle (Molecular Events): The Powerhouse
The cross-bridge cycle is a blindingly fast, repetitive, four-step biochemical engine sequence that physically causes the thin filaments to slide over the thick filaments.
Cross-Bridge Formation (The Latch)
The energized, upright ("cocked") myosin head—which is already tightly holding onto a spent ADP and inorganic phosphate (Pi) molecule from the previous cycle—possesses a massive chemical attraction (affinity) for the exposed actin filament. The moment the tropomyosin shifts to uncover the binding sites (thanks to calcium), the myosin head violently snaps upward and forms a strong, unbreakable physical link with the actin. This physical connection between thick and thin filaments is the famous "cross-bridge."
The Power Stroke (The Pull)
The instant physical formation of the cross-bridge chemically triggers the immediate release of the trapped inorganic phosphate (Pi) from the myosin head. This release unleashes the stored mechanical energy, causing the myosin head to forcefully pivot on its hinge, jerking from its high-energy 90° angle to a low-energy, bent 45° angle. This brutal pivoting movement is the power stroke. Because it is firmly attached like a grappling hook, the pivoting myosin head violently drags the entire thin actin filament a short microscopic distance (~10 nm) toward the dead center (M-line) of the sarcomere. Immediately following the pivot, the remaining ADP molecule is ejected, leaving the myosin head in a rigid, low-energy state, still tightly locked onto the actin.
Cross-Bridge Detachment (The Release)
Following the power stroke, the spent myosin head remains rigidly "stuck" to the actin in a low-energy configuration (known as the "rigor" state). The absolute only biochemical way for the myosin head to let go of the actin is for a brand new, fresh molecule of ATP to physically bind to the empty ATP-binding site on the back of the myosin head. The instant this new ATP binds, it causes an allosteric (shape) change that severely weakens the molecular bond between the myosin and the actin, reducing their chemical affinity to zero, and forcing the myosin head to abruptly detach.
Re-cocking of the Myosin Head (The Reset)
The completely detached myosin head, now carrying its fresh ATP, immediately acts as an aggressive enzyme (myosin ATPase). It instantly hydrolyzes (burns) the ATP, snapping it into ADP and inorganic phosphate (Pi). The explosive energy released from breaking this intense ATP chemical bond is captured entirely by the myosin hinge, physically forcing the head to snap backward, moving from its low-energy bent position back to its high-energy, upright, "cocked" position. It is now fully energized, reset, and coiled like a rat-trap, ready to violently repeat the entire cycle by latching onto another active site further down the actin filament (provided Ca²⁺ is still present in the sarcoplasm keeping the sites exposed).
Clinical Insight: Rigor Mortis
Why do bodies become stiff after death? Upon death, cellular respiration halts, and ATP production completely drops to zero. Simultaneously, cell membranes degrade, allowing calcium to leak into the sarcoplasm, triggering Step 1 (Cross-Bridge Formation) and Step 2 (The Power Stroke). However, because there is absolutely no new ATP being produced by the dead body, Step 3 (Detachment) cannot occur. Millions of myosin heads remain permanently, rigidly locked onto the actin filaments in the low-energy rigor state. The entire muscular system locks into a solid, unbreakable spasm known as rigor mortis, which only subsides days later when the muscle proteins literally begin to rot and decompose.
3. Sarcomere Shortening: The Ultimate Result
As millions of myosin heads asynchronously paddle through this cycle thousands of times per second, the results are profound across the geometry of the cell:
- The thin filaments actively slide inward, aggressively pulled past the stationary thick filaments toward the M-line.
- The Z-discs on either end of the sarcomere are violently dragged closer together, physically shortening the entire microscopic sarcomere unit.
- Under a microscope, the light I bands and the central H zone compress and shorten.
- The dark A band remains absolutely unchanged in length, proving the thick filaments do not shrink.
When hundreds of thousands of sarcomeres lined up in series shorten simultaneously, the entire gross muscle belly shortens, pulling the tendon and generating massive skeletal force.
IX. Muscle Relaxation
Muscle relaxation is not simply a passive fading out; it is a highly active, metabolically expensive, energy-requiring biochemical process. You must burn ATP to relax!
- Cessation of Motor Neuron Signal: The upper brain commands stop, the lower motor neuron halts its action potential firing, and no new ACh is exocytosed into the NMJ cleft.
- AChE Activity: The remaining ACh currently sitting in the synaptic cleft is rapidly and aggressively broken down into choline and acetate by the ever-present enzyme acetylcholinesterase, completely starving the nAChR receptors of their ligand.
- Repolarization of Sarcolemma: The muscle fiber nAChR channels slam shut, Na⁺ influx ceases, and the sarcolemma and deep T-tubules fully repolarize back to a negative resting state.
- Ca²⁺ Reuptake into SR (The Heavy Lifting): As the T-tubules repolarize, the DHPR voltage sensors return to normal, pushing the massive RyR calcium release channels on the SR closed. Simultaneously, thousands of active transport vacuums called SERCA pumps (Sarcoplasmic/Endoplasmic Reticulum Calcium ATPase) burn massive amounts of ATP to aggressively pump the escaped Ca²⁺ ions from the sarcoplasm backward, against their gradient, into the SR storage vaults.
- Tropomyosin Blocks Active Sites: As the ruthless SERCA pumps drop the ambient sarcoplasmic Ca²⁺ levels back to near zero, calcium physically detaches from the Troponin C receptor. The Troponin complex snaps back to its original, un-calcium-bound shape. This shape change allows the long tropomyosin rod to physically shift back over, snapping into place and completely re-covering and blocking the myosin-binding sites on the actin filament.
- Muscle Relaxes: With the binding sites completely shielded, no new cross-bridges can possibly form. The remaining attached myosin heads finish their current cycle, detach, and are physically barred from reattaching. The muscle fiber loses tension and passively lengthens back to its resting resting length due to gravity or the pull of an opposing antagonist muscle.
Malignant Hyperthermia
This is a rare, life-threatening genetic emergency triggered by the administration of certain general anesthetics (like halothane) or muscle relaxants (like succinylcholine) during surgery. The patient possesses a mutated, defective Ryanodine Receptor (RyR). When exposed to the anesthetic gas, the mutant RyR jams wide open, dumping an endless, unstoppable flood of calcium into the sarcoplasm. The muscles instantly go into a massive, full-body hyper-metabolic contraction. The SERCA pumps burn through the body's entire ATP supply trying to pump the calcium back in, generating highly lethal amounts of bodily heat (hyperthermia) and lactic acid. Without immediate administration of the antidote Dantrolene (which specifically acts to forcefully slam the RyR channels shut), the patient will rapidly die of a metabolic meltdown on the operating table.
X. List of References
This comprehensive synthesis of excitable tissue physiology is got from and aligns with the following gold-standard academic texts and resources universally utilized in medical and physiological education:
- Hall, J. E., & Guyton, A. C. (2015). Guyton and Hall Textbook of Medical Physiology (13th ed.). Philadelphia, PA: Elsevier. (Primary source for comprehensive AP mechanics and excitation-contraction coupling).
- Costanzo, L. S. (2018). Physiology (6th ed.). Philadelphia, PA: Elsevier. (Primary source for succinct, step-by-step molecular breakdowns of the cross-bridge cycle and NMJ pharmacology).
- Boron, W. F., & Boulpaep, E. L. (2016). Medical Physiology (3rd ed.). Philadelphia, PA: Elsevier. (Advanced reference for SNARE protein mechanics, spatial/temporal summation mathematical integration, and specific ion channel gating kinetics).
- Katzung, B. G., Masters, S. B., & Trevor, A. J. (2021). Basic & Clinical Pharmacology (15th ed.). McGraw-Hill Education. (Reference for applied clinical correlations, including Botox, local anesthetics, and Malignant Hyperthermia antidotes).