
PHYSIOLOGY OF EXCITABLE TISSUES
By the end of this exhaustive physiological guide, you will master:
- The fundamental definition of Excitability and the specialized cells that possess it.
- The electrochemical mechanisms maintaining the Resting Membrane Potential (RMP).
- The exact structural and functional gating mechanisms of Ion Channels.
- The step-by-step ionic basis of the Action Potential and signal propagation.
- The mechanisms of Inhibition (hyperpolarization, presynaptic, and pharmacological).
- The profound Clinical Significance of excitability, focusing heavily on electrolyte imbalances and neurological diseases.
1. Introduction to Excitability
Excitability refers to the fundamental ability of a living cell to respond to an environmental stimulus by generating a rapid, highly coordinated electrical signal known as an Action Potential. It is defined as a measurable physical and chemical change that occurs across the cell membrane when a stimulus is applied to a tissue. A stimulus is any external or internal agent (electrical, chemical, thermal, or mechanical) that produces this excitation.
The action potential is a transient, rapid, and self-propagating reversal of the electrical potential across the cell membrane. This electrical signal is the medium through which cells rapidly transmit information, either along the length of an individual cell (like a wire) or to other cells via specialized junctions (synapses). This property is crucial for rapid communication and coordination within the body, underpinning virtually every complex physiological function, from perception, thought, and memory to voluntary movement and visceral regulation.
The Electrical Tripwire
Think of an excitable cell like a highly sensitive electrical tripwire or alarm system.
- The Resting State is the armed system, charged and waiting for a trigger.
- The Stimulus is the physical pressure that activates the tripwire.
- The Action Potential is the immediate, swift, and uniform "alarm bell" that rings loudly and clearly, sending its message undiminished through the entire system to orchestrate a massive, coordinated response.
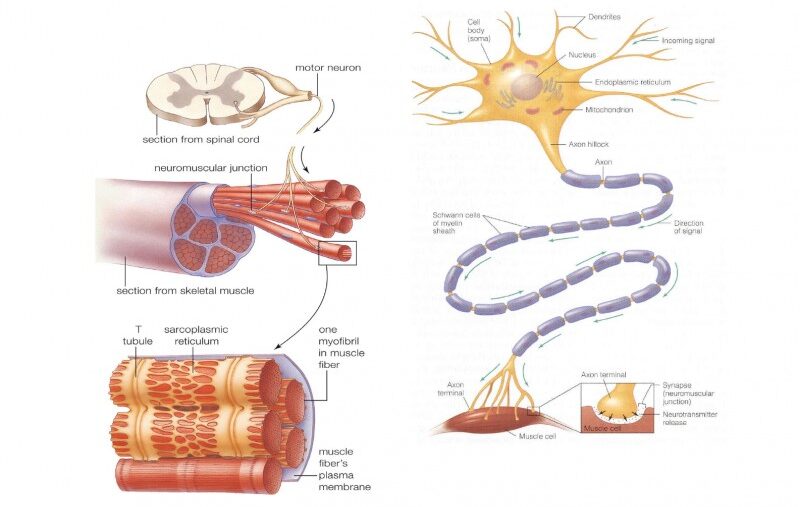
2. Types of Excitable Cells
While all living cells exhibit some degree of responsiveness to their environment, only a select group possess the highly specialized membrane proteins and machinery required to generate and propagate rapid electrical action potentials. These are the "excitable cells."
The Master Communicators
Expanded Role: Neurons are the fundamental functional units of the nervous system. Their primary function is the rapid transmission of electrical and chemical signals for sensory input, integration, motor output, cognition, and emotion.
Unique Features: They possess specialized anatomical structures: dendrites (the "antennae" to receive signals), a cell body (soma) (the metabolic center), and a long axon (the "transmission cable" to send signals), which is often insulated by a lipid-rich myelin sheath to drastically speed up conduction velocity.
The Effectors of Movement
Muscle cells are specialized for contraction, which generates physical force and movement. Their electrical excitability is the absolute prerequisite for this mechanical action (Excitation-Contraction Coupling).
- Skeletal Muscle Cells: Responsible for all voluntary movements (walking, speaking, breathing). When a somatic motor neuron sends an action potential, it triggers a massive muscle action potential across the sarcolemma, leading to instantaneous contraction.
- Cardiac Muscle Cells: Found only in the myocardium, responsible for the rhythmic and involuntary pumping of blood. They possess autorhythmicity (can generate their own pace) and have distinctively long action potentials featuring a "plateau phase" to ensure coordinated, non-tetanic contractions.
- Smooth Muscle Cells: Mediate involuntary movements in the walls of internal organs like the digestive tract, blood vessels, and urinary bladder. Their excitability is influenced heavily by stretch, local chemical mediators, and the autonomic nervous system.
The Secretory Responders
Expanded Role: Many glandular cells (e.g., in the adrenal medulla, pancreatic beta-cells) exhibit true excitability. They can respond to an electrical stimulus from a neuron or a chemical change (like high blood glucose) by generating their own electrical depolarization event.
Excitability Link: This electrical event is typically coupled directly to the release of their internal secretions (hormones, digestive enzymes). For example, adrenal medullary cells depolarize in response to a sympathetic neuronal signal, triggering massive Calcium (Ca²+) influx and the immediate exocytosis of epinephrine. This ensures precise, rapid control over systemic hormone release.
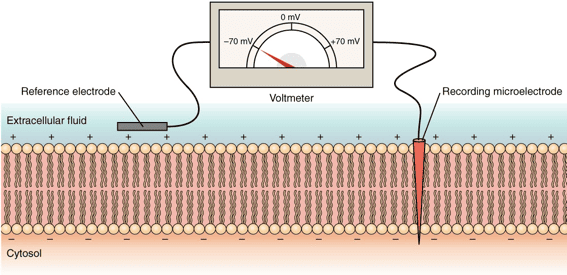
3. The Membrane Potential
The capacity of these cells to generate electrical signals rests entirely on the foundational concept of the Membrane Potential. This is the voltage difference across the cell's outer boundary (the lipid bilayer). It represents stored, potential electrical energy created by an uneven distribution of ions (electrically charged particles) between the Intracellular Fluid (ICF) and the Extracellular Fluid (ECF).
Resting Membrane Potential (RMP)
When an excitable cell is quiet, it maintains a stable, baseline electrical charge called the Resting Membrane Potential (RMP). In this state, the inside of the cell consistently holds a negative charge relative to the outside. Standard RMP values include:
- Neurons: Typically around -70 mV.
- Skeletal & Cardiac Muscle: Typically around -90 mV.
Creating and Maintaining the RMP
The RMP is not static; it is a highly dynamic state, constantly maintained by an active, energy-consuming interplay of three major factors:
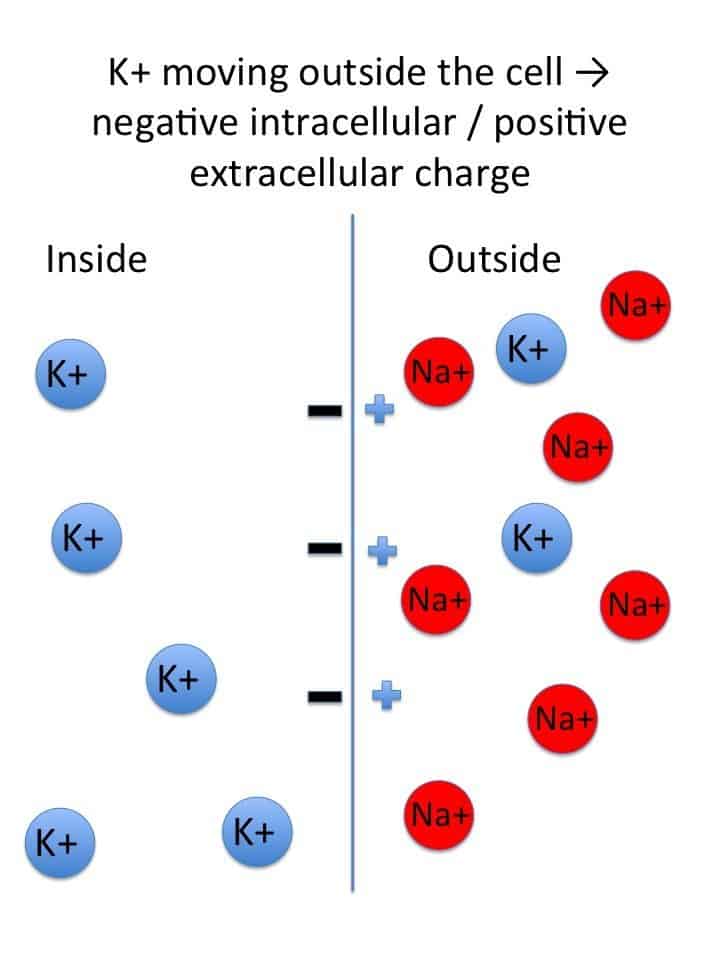
- Ion Gradients (The Concentration Divide): The foundational requirement is the extreme difference in the concentrations of key ions: a massive concentration of Sodium (Na+) outside the cell and a massive concentration of Potassium (K+) inside the cell.
- Selective Permeability (The Leaky Gates): At rest, the cell membrane is 50 to 100 times more permeable to K+ than to Na+. This is because the membrane possesses many more open K+ "leak" channels than Na+ leak channels. Consequently, K+ constantly leaks out of the cell, carrying positive charge away and leaving the inside of the cell negative.
- Sodium-Potassium ATPase Pump (The Gradient Upholder): This ubiquitous active transporter continually burns cellular energy (ATP) to pump 3 Na+ ions OUT for every 2 K+ ions it pumps IN. This directly maintains the steep concentration gradients and, because it removes more positive charge than it brings in, it contributes a small amount to the RMP's negativity (acting as an electrogenic pump).
Deeper: Equilibrium Potential
The Nernst Equation
The Equilibrium Potential for a specific single ion is the exact membrane voltage at which there is no net movement of that ion across the membrane. At this specific voltage, the electrical force pulling the ion in one direction is perfectly, equally balanced by the chemical (concentration) force pushing it in the other direction. The Nernst Equation calculates this theoretical value:
Eion = (RT / zF) * ln([ion]out / [ion]in)
Physiological Context: The equilibrium potential for Potassium (EK) is approximately -90 mV. The equilibrium potential for Sodium (ENa) is approximately +60 mV. Because the resting membrane is highly permeable to Potassium and mostly impermeable to Sodium, the overall RMP (-70 mV) sits very close to the Potassium equilibrium potential!
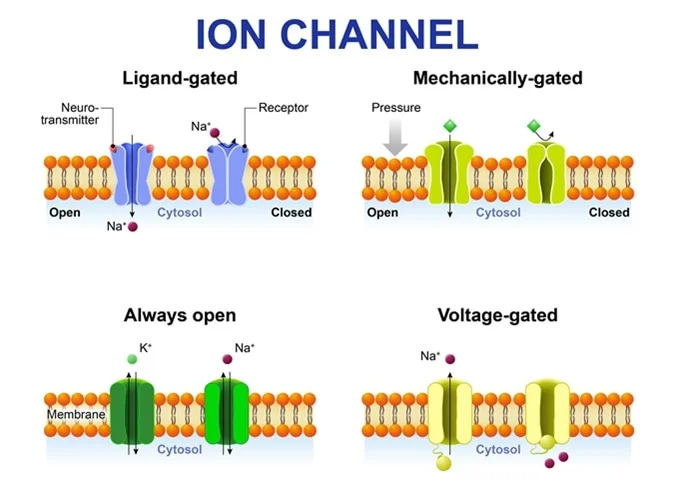
4. Ion Channels: The Gates of Excitability
Ion channels are highly specialized, complex transmembrane proteins that form selective pores allowing specific ions to cross the otherwise impermeable lipid bilayer.
Types Relevant to Excitability:
1. Leak Channels (Non-gated)
These channels are essentially always open. They are instrumental in establishing the RMP, particularly the widespread K+ leak channels that allow the continuous efflux of positive charge.
2. Gated Channels (The Responsive Switches)
These channels have molecular "gates" that open or close only in response to a particular physiological trigger. They are the absolute essential machinery for generating action potentials.
| Type of Gated Channel | Trigger Mechanism | Physiological Example |
|---|---|---|
| Voltage-Gated Channels | Open or close in direct response to changes in the transmembrane voltage. They possess charged amino acids that physically move when the voltage changes. | Voltage-gated Na+ channels (driver of depolarization) and Voltage-gated K+ channels (driver of repolarization). |
| Ligand-Gated Channels (Chemically Gated) | Open or close only when a specific chemical messenger (a ligand), such as a neurotransmitter or hormone, binds to a receptor site on the channel. | The Nicotinic Acetylcholine Receptor at the neuromuscular junction. Binding of ACh opens the pore to let Na+ rush in. |
| Mechanically Gated Channels | Open or close when the membrane itself is physically deformed, stretched, or subjected to pressure. | Pacinian corpuscles in the skin (deep pressure receptors) or hair cells in the inner ear (responding to sound waves). |
5. Initiating the Response: Stimulus and Threshold
The Stimulus: A Call to Action
A stimulus is any detectable change (electrical, chemical, or mechanical) in the cell's environment that has the potential to alter its resting membrane potential. Stimuli can cause two types of local changes:
- Depolarization: A shift in membrane voltage where the inside of the cell becomes less negative (e.g., moving from -70 mV up to -50 mV). This is an excitatory shift.
- Hyperpolarization: A shift where the inside of the cell becomes more negative (e.g., moving from -70 mV down to -90 mV). This is an inhibitory shift.
Threshold: The Point of No Return
Threshold is the critical, specific voltage level that a depolarization must reach for an action potential to fire (typically around -55 mV in neurons). It operates on an "all-or-none" principle: if a stimulus causes a local depolarization that reaches this threshold, a full, unstoppable action potential fires. If the stimulus is weak and only depolarizes the cell to -60 mV, nothing happens, and the potential simply dissipates.
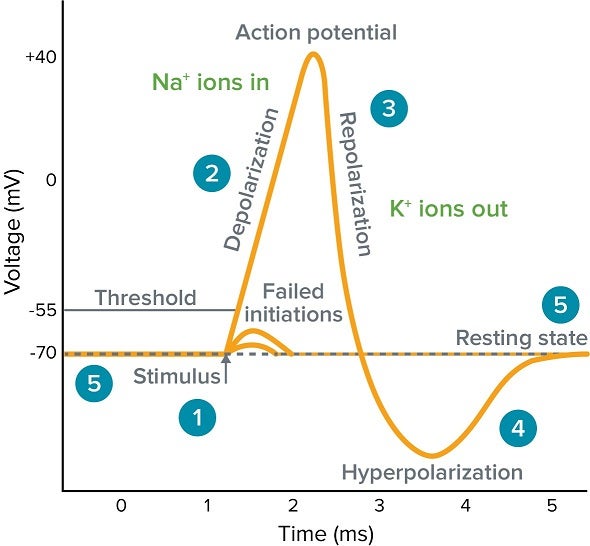
6. The Action Potential: Step-by-Step
The action potential is the primary electrical signal employed by excitable cells to swiftly transmit information across massive anatomical distances (like from the spinal cord to the toe). It stands as an "all-or-nothing" phenomenon: once initiated, it proceeds through its entire sequence with consistent strength, never diminishing over distance.
| Stage | Membrane Potential | Ionic Movement & Channel Status |
|---|---|---|
| 1. Resting State | -70 mV | All voltage-gated Na+ and K+ channels are CLOSED. The RMP is maintained by K+ leak channels and the active Na+/K+ pump. |
| 2. Depolarization to Threshold | -70 mV to -55 mV | A local stimulus causes a few voltage-gated Na+ channels to open. A small amount of Na+ trickles into the cell. If enough Na+ enters to push the voltage to the -55 mV threshold, the cell commits to firing. |
| 3. Rising Phase (Depolarization) | -55 mV to +30 mV | Threshold triggers a massive positive feedback loop. A vast number of voltage-gated Na+ channels rip open. A massive and swift surge of Na+ rushes into the cell, causing the inside to rapidly become positive (+30 mV). |
| 4. Repolarization Phase | +30 mV back down toward -70 mV | At the +30 mV peak, the voltage-gated Na+ channels inactivate (a physical "plug" blocks the pore), instantly stopping Na+ influx. Simultaneously, the slower voltage-gated K+ channels finally open fully. A massive outflow of K+ rapidly restores the membrane's negative internal charge. |
| 5. Afterhyperpolarization (Undershoot) | Dips below -70 mV (e.g., -80 mV) | The voltage-gated K+ channels are sluggish and close slowly. K+ continues to exit for a brief period, causing the membrane to become temporarily more negative than the baseline RMP. |
| 6. Return to Rest | Returns to -70 mV | The slow K+ channels finally close completely. The ever-active Na+/K+ pump and normal leak channels re-establish the original resting ion concentration gradients. |
Defining Features of Action Potentials
- All-or-Nothing: If the threshold is crossed, the action potential unfolds completely with the exact same magnitude and shape every time. If threshold is not reached, no action potential occurs.
- Non-Decremental: Action potentials are continuously re-generated along the membrane. A signal starting in the brain is just as strong when it reaches the foot.
7. Refractory Periods & Signal Propagation
Refractory Periods (The Reset Time)
A cell cannot fire continuously without resetting. This required downtime is called the refractory period.
- Absolute Refractory Period: Occurs during the entire rising phase and most of the repolarization phase. During this time, the voltage-gated Na+ channels are either already fully open or rigidly inactivated. Therefore, absolutely NO second stimulus, regardless of how massively strong it is, can trigger another action potential. Function: This ensures one-way propagation of the signal and prevents the signal from traveling backward.
- Relative Refractory Period: Occurs during the afterhyperpolarization phase. The Na+ channels have reset to their closed (but ready) state, but the cell is hyperpolarized (extra negative). A stimulus can provoke another action potential, but it must be significantly stronger than normal to overcome the hyperpolarization.
Propagation of Action Potentials: Spreading the Message
The massive electrical shift at one point on the membrane triggers the opening of voltage-gated Na+ channels in the immediately adjacent area. This process repeats endlessly, moving the signal down the length of the nerve or muscle fiber like a burning fuse.
Myelination: Enhancing Speed via Saltatory Conduction
Many nerve fibers are thickly insulated by a fatty lipid layer called the Myelin Sheath (produced by Schwann cells in the PNS and Oligodendrocytes in the CNS). Action potentials cannot form where myelin exists. Therefore, the signal must "jump" from one microscopic uninsulated gap (a Node of Ranvier) to the next. This rapid, jumping propagation is termed Saltatory Conduction and dramatically increases the signal's speed while saving vast amounts of cellular energy.
Factors Influencing Conduction Speed:
- Fiber Diameter: Larger diameter fibers conduct signals much more quickly because there is less internal electrical resistance.
- Myelination: Myelinated fibers transmit signals up to 50 times faster than unmyelinated fibers.
8. Inhibition of Excitability
Just as cells must generate signals to act, they desperately need ways to inhibit signals, ensuring precise motor control, emotional regulation, and preventing uncontrolled, chaotic firing (seizures).
Driving Further from Threshold
Inhibitory neurotransmitters (like GABA in the brain or Glycine in the spinal cord) bind to ligand-gated channels that selectively allow Cl− (Chloride) to enter the cell or K+ to leave. The outcome is an immediate increase in the negative charge inside the cell (e.g., dropping from -70 mV to -80 mV). This creates an Inhibitory Postsynaptic Potential (IPSP), making it significantly harder for any excitatory stimulus to push the cell to threshold.
Muting the Signal at its Source
An inhibitory neuron releases a neurotransmitter (e.g., GABA) directly onto the axon terminal of an excitatory neuron. This slightly reduces the electrical charge of the terminal. Consequently, when an action potential arrives at that terminal, fewer voltage-gated Calcium channels open, meaning fewer excitatory neurotransmitters are released into the synaptic cleft. This allows for brilliant fine-tuning and selective dampening of specific signals without shutting down the whole system.
Manipulating Channels with Drugs
A vast array of modern drugs and natural toxins work by directly interfering with ion channels to deliberately shut down excitability.
- Local Anesthetics (e.g., Lidocaine): Physically plug the intracellular pore of voltage-gated Na+ channels. Without Na+ influx, pain-sensing nerves cannot generate action potentials, resulting in numbness.
- Tetrodotoxin (TTX): A highly potent neurotoxin found in pufferfish. It permanently blocks the extracellular side of voltage-gated Na+ channels, causing rapid, fatal respiratory paralysis.
- Anti-epileptic Drugs (e.g., Phenytoin): Work by stabilizing Na+ channels in their inactivated state, preventing the excessively rapid, repetitive firing seen in seizures.
9. Clinical Significance of Excitability
An in-depth comprehension of cellular excitability is absolutely vital for understanding, diagnosing, and creating effective medical treatments for numerous devastating conditions affecting the nervous system and musculature.
Conditions of the Nervous System and Muscles:
- Epilepsy: Marked by episodes of abnormal, highly synchronized, and excessive electrical firing of large groups of neurons in the cerebral cortex, resulting in seizures. It represents a catastrophic failure of normal inhibitory mechanisms.
- Multiple Sclerosis (MS): A severe autoimmune disease where the myelin sheath insulating central nerve fibers is destroyed by the body's own immune system. This exposes the underlying axon, drastically slowing, weakening, or completely blocking action potential propagation. It leads to progressive muscle weakness, blindness, and sensory disturbances.
- Myasthenia Gravis: An autoimmune disease that generates antibodies to destroy Nicotinic acetylcholine receptors at the neuromuscular junction. This reduces the ability of motor nerve signals to adequately excite muscle cells, leading to profound, easily fatigued muscle weakness (ptosis, difficulty breathing).
- Cardiac Arrhythmias: Irregular heart rhythms stemming from abnormalities in the electrical excitability, automaticity, or conduction pathways of heart muscle cells. This leads to potentially fatal disruptions to the heart's synchronized pumping action (e.g., Ventricular Fibrillation).
10. The Critical Role of Electrolyte Imbalances
Because the entire system of excitability relies on precise concentration gradients of ions, systemic electrolyte imbalances alter the very foundation of the resting membrane potential and threshold, leading to severe clinical emergencies.
| Electrolyte Imbalance | Pathophysiological Mechanism & Cellular Effect | Clinical Consequences |
|---|---|---|
| Hyperkalemia (Elevated K+) |
High extracellular K+ prevents K+ from leaking out of the cell. The resting membrane potential becomes less negative (e.g., moves from -90mV to -70mV in the heart), putting it dangerously close to threshold. While this initially causes twitchy hyperexcitability, prolonged depolarization permanently inactivates voltage-gated Na+ channels, ultimately rendering cells completely inexcitable. | Lethal cardiac arrhythmias, characteristic "peaked T-waves" on an ECG, and eventual flaccid paralysis leading to cardiac arrest. |
| Hypokalemia (Low K+) |
Low extracellular K+ creates a steeper gradient, forcing too much K+ to leak out of the cell. The RMP becomes more negative (hyperpolarized, e.g., dropping to -100mV). This moves the cell much further away from the threshold, making it significantly harder to fire an action potential. | Profound muscle weakness, fatigue, diminished reflexes, and dangerous heart arrhythmias (prominent U-waves on ECG). |
| Hyponatremia / Hypernatremia (Sodium Imbalances) |
Because the rapid, massive influx of Na+ is the absolute primary driver of the depolarization spike, severe imbalances in ECF Na+ levels disrupt the amplitude (height) and speed of action potentials. Furthermore, severe sodium imbalances cause massive cellular swelling or shrinking due to osmotic shifts. | Confusion, lethargy, severe muscle twitching, seizures, coma, and brain damage. |
| Hypocalcemia (Low Ca²+) |
Paradoxical Effect: Calcium ions normally bind to the outside of the cell membrane, physically "shielding" Na+ channels. When calcium is low, this shield is removed. Voltage-gated Na+ channels become highly unstable and rip open at much lower, more negative voltages. The threshold is dangerously lowered. | Severe, involuntary muscle spasms, cramps, and tetany (e.g., Trousseau's sign and Chvostek's sign). |
| Hypercalcemia (High Ca²+) |
High extracellular calcium heavily coats the membrane, stabilizing the Na+ channels and making them incredibly stiff and hard to open. The threshold is raised, drastically decreasing neuronal excitability. | "Bones, stones, groans, and psychiatric overtones." Severe muscle weakness, lethargy, depressed reflexes, and reduced overall neurological function. |
References
- Guyton and Hall: Textbook of Medical Physiology. (Chapters on Membrane Potentials, Action Potentials, and Contraction of Skeletal Muscle).
- Katzung, B. G.: Basic & Clinical Pharmacology. (Chapters on Local Anesthetics and Drugs Acting on the Central Nervous System).
- Ganong, W. F.: Review of Medical Physiology. (Excitable Tissue: Nerve & Muscle).
- Costanzo, L. S.: Physiology. (Cellular Physiology: Resting Membrane Potential and Action Potentials).