Introduction to Basic Pharmacology
Introduction to Basic Pharmacology and Pharmacology Practicals (Instrumentation)
Learning Outcomes of the Lecture
By the end of this comprehensive guide, students should be fully equipped to:
- Define pharmacology and clearly outline its major branches and sub-disciplines.
- Distinguish definitively between the concepts of pharmacodynamics and pharmacokinetics.
- Explain the critical importance of pharmacology practicals and laboratory experiments in medical and scientific training.
- Identify and understand the major instruments and equipment used in modern and historical pharmacology laboratories.
- Describe the specific functions of organ bath systems, transducers, and recording devices in depth.
- Recognize and apply ethical considerations (such as the 3Rs) in experimental pharmacology.
Introduction to Pharmacology
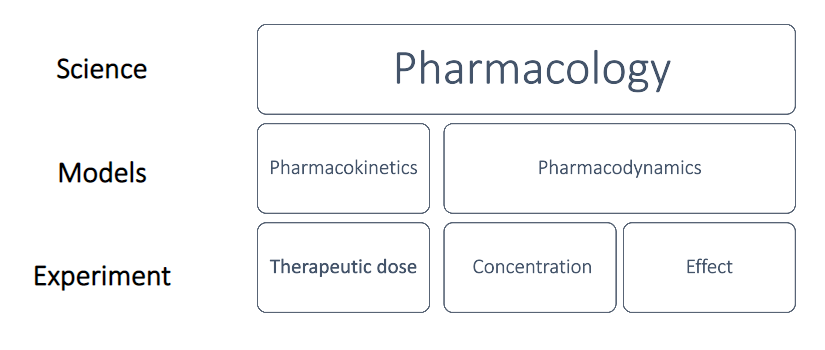
Pharmacology is broadly defined as the rigorous scientific study of drugs and their interactions with living systems. Derived from the Greek words pharmakon (drug or poison) and logos (study), it is a vast field that examines every aspect of how drugs produce their physiological effects, how the human (or animal) body processes these foreign substances, and how these chemicals can be utilized therapeutically to treat disease, or experimentally to understand biological processes.
A drug, in the context of pharmacology, can be defined as any chemical substance (natural, synthetic, or endogenous) that modifies physiological or biochemical functions when administered to a living organism. This includes everything from life-saving antibiotics to everyday pain relievers, as well as substances of abuse and environmental toxins.
The Multidisciplinary Nature of Pharmacology
Pharmacology does not exist in isolation. It acts as a bridge between the physical sciences and the biological sciences. It integrates core knowledge from several crucial disciplines, including:
- Physiology: Understanding normal body functions is essential before one can understand how a drug alters those functions.
- Biochemistry: Provides the foundation for understanding the chemical basis of drug action at the enzymatic and metabolic levels.
- Molecular Biology: Helps in understanding how drugs interact with genetic material, intracellular signaling, and protein synthesis.
- Medicinal Chemistry: Focuses on the structural design, synthesis, and optimization of pharmaceutical drugs.
- Toxicology: The study of the adverse or poisonous effects of chemicals, closely tied to drug safety.
- Clinical Medicine: The ultimate application of pharmacological knowledge to diagnose, prevent, and treat illnesses in human patients.
Major Branches of Pharmacology
To fully grasp pharmacology, the field is traditionally divided into several distinct, yet deeply interconnected, branches.
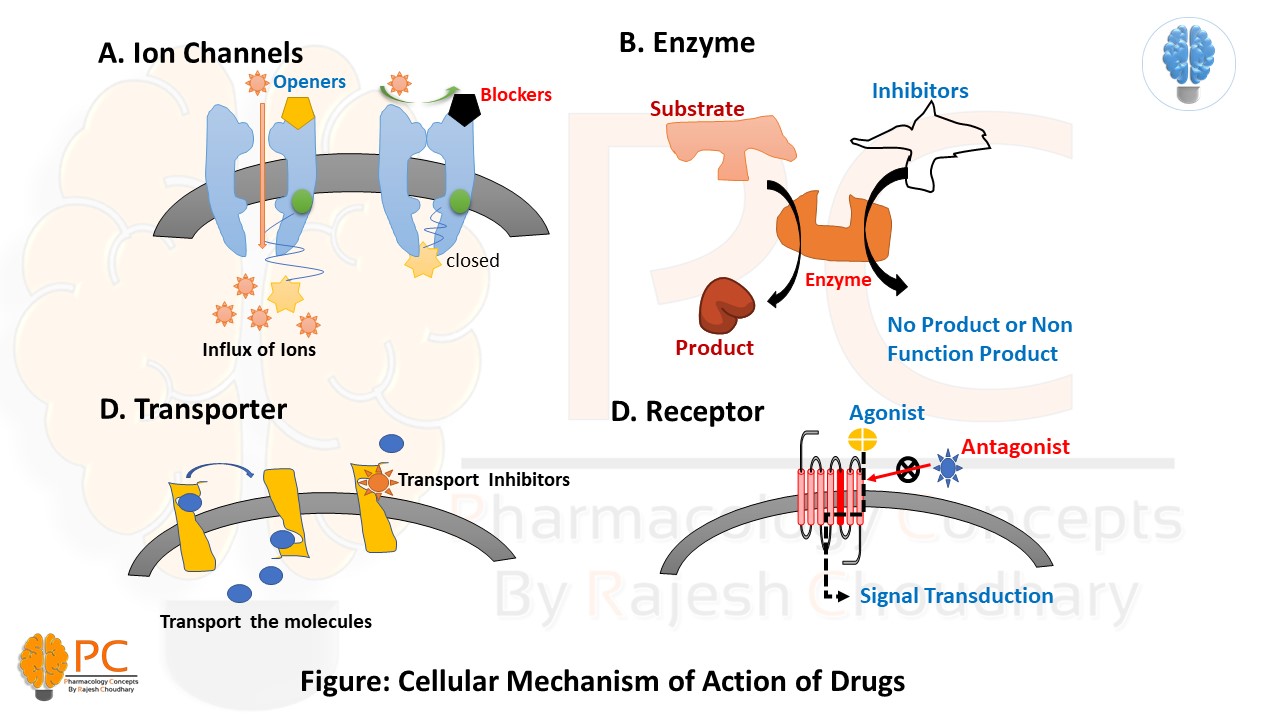
1. Pharmacodynamics
Pharmacodynamics essentially studies what the drug does to the body. It delves into the specific biochemical and physiological effects of drugs and their mechanisms of action.
Key aspects of pharmacodynamics include:
- Mechanism of Drug Action: Exactly how a drug produces its effect at the cellular level.
- Drug–Receptor Interactions: How drugs bind to specific protein targets (receptors) to initiate or block a biological response.
- Cellular Targets: Drugs typically exert their effects by interacting with four main regulatory proteins:
- Ion Channels: Drugs can act as openers (increasing ion influx) or blockers (preventing ion passage).
- Enzymes: Drugs often act as inhibitors, preventing the enzyme from converting a substrate into a product (e.g., aspirin inhibiting COX enzymes).
- Transporters: Drugs can act as transport inhibitors, preventing the movement of molecules across cell membranes.
- Receptors: Drugs can be Agonists (activating the receptor for signal transduction) or Antagonists (blocking the receptor and preventing activation).
- Dose–Response Relationships: The mathematical and graphical relationship between the amount of drug given (dose) and the magnitude of the effect produced. As the log of the drug concentration increases, the effect typically increases until a maximum plateau is reached.
- Therapeutic and Toxic Effects: Determining the primary intended effects versus unintended side effects.
How β-blockers reduce heart rate: A beta-blocker (like Atenolol) acts as an antagonist. It specifically targets and blocks β1-adrenergic receptors located in the heart muscle. By blocking these receptors, it prevents adrenaline from binding, which structurally and functionally reduces the heart rate and blood pressure (this is what the drug does to the body).
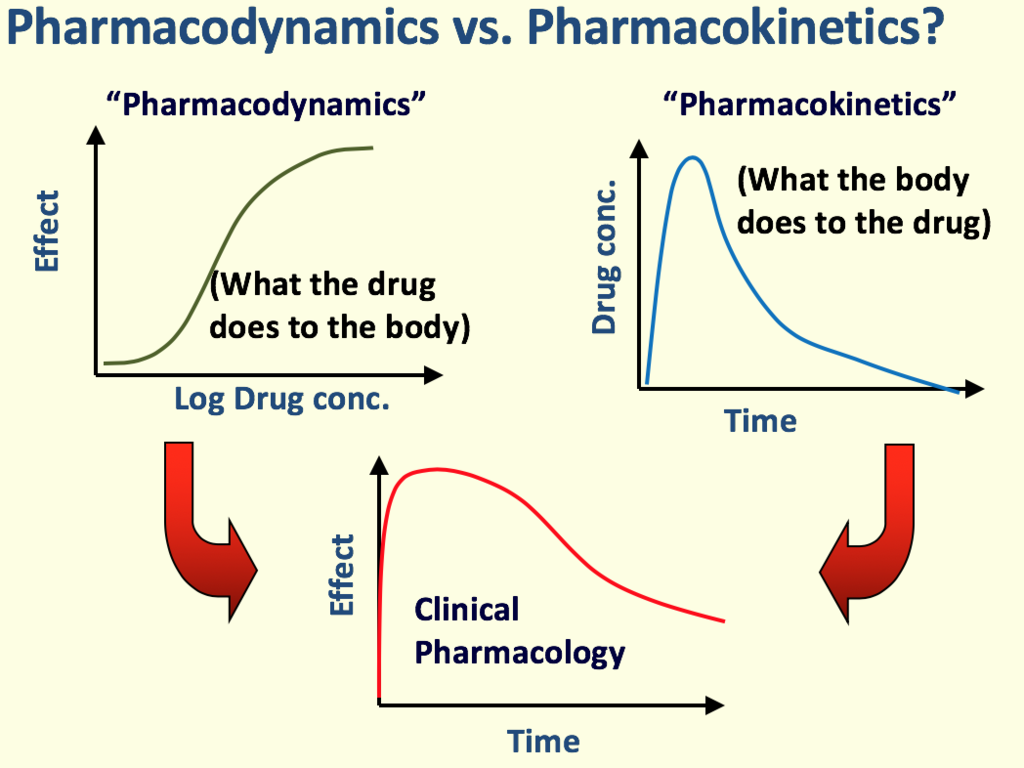
2. Pharmacokinetics
Pharmacokinetics studies what the body does to the drug. It traces the journey of a drug molecule from the moment it enters the body until it is completely removed.
It involves four major, continuous processes, universally remembered by the acronym ADME:
The movement of a drug from its site of administration (e.g., gut, muscle, skin) into the systemic blood circulation. Factors like route of administration, lipid solubility, and pH heavily influence this.
The reversible transfer of a drug from one location to another within the body, typically from the bloodstream into tissues, organs, and intracellular spaces. It is affected by blood flow, tissue binding, and membrane permeability (e.g., the blood-brain barrier).
The chemical modification or breakdown of drugs, primarily occurring in the liver. The body attempts to make the drug more water-soluble so it can be easily excreted.
The irreversible elimination of the drug and its metabolites from the body. The kidneys (via urine) are the primary route, but drugs can also be excreted through bile, feces, sweat, saliva, tears, and lungs (exhaled air).
First-pass metabolism of drugs like propranolol: When propranolol is taken orally, it is absorbed by the digestive tract and carried directly to the liver via the hepatic portal vein. The liver highly metabolizes (destroys) a large portion of the drug before it ever reaches the systemic circulation. This "first-pass effect" drastically reduces the bioavailability of the drug, which is an example of what the body does to the drug.
3. Therapeutics (Clinical Pharmacology/Pharmacotherapeutics)
This branch focuses strictly on the clinical use of drugs to prevent, diagnose, or treat diseases. It is the practical application of pharmacology in a healthcare setting, emphasizing evidence-based medicine, rational prescribing, and patient care.
- Antihypertensive therapy: Using drugs to lower high blood pressure and prevent cardiovascular events.
- Antidiabetic therapy: Managing blood sugar levels using insulin or oral hypoglycemic agents.
- Antimicrobial therapy: Utilizing antibiotics, antivirals, or antifungals to eradicate infections while minimizing harm to the host.
4. Toxicology
Toxicology is the study of the harmful, adverse, or toxic effects of drugs, chemicals, and environmental poisons on living systems. Paracelsus famously stated, "The dose makes the poison," highlighting that any drug can be toxic if taken in excess.
It includes the study of:
- Acute toxicity: Harmful effects occurring rapidly after a single or short-term exposure.
- Chronic toxicity: Harmful effects resulting from prolonged, long-term repeated exposure.
- Organ-specific toxicity: Such as hepatotoxicity (liver damage), nephrotoxicity (kidney damage), or cardiotoxicity (heart damage).
- Poison management: The clinical strategies to treat overdoses, including the administration of specific antidotes.
5. Experimental Pharmacology
This branch studies drug effects under strictly controlled laboratory conditions using various experimental models. It forms the crucial foundation for the entire pharmaceutical industry's drug discovery pipeline and preclinical testing phases (before a drug is ever tested in humans).
Models include:
- Isolated tissues: Organs or tissues removed from an animal and kept alive in nutrient solutions (e.g., isolated heart, intestine).
- Laboratory animals: Whole living organisms (in vivo studies), usually rodents like mice, rats, or guinea pigs, to observe systemic effects.
- Cellular models: Cultured human or animal cells grown in petri dishes (in vitro studies).
- Molecular assays: Biochemical tests to observe drug-target interactions at the genetic or protein level.
The Importance of Pharmacology Practicals
Theoretical knowledge alone is insufficient for scientific mastery. Pharmacology practicals (laboratory sessions) are a cornerstone of medical and scientific curricula. They serve to bridge the gap between textbook theories and real-world biological phenomena.
Practicals help students and researchers to:
- Understand drug actions experimentally: Seeing a physical tissue respond to a drug solidifies abstract concepts.
- Learn fundamental research techniques: Mastering the use of delicate instruments, precise pipetting, and tissue handling.
- Develop skills in experimental design: Learning how to formulate hypotheses, set up controls, and execute a valid scientific test.
- Interpret dose-response relationships: Practically gathering data points to plot logarithmic curves and calculate metrics like ED50 (Effective Dose 50%).
- Understand biological variability: Recognizing that living tissues do not behave like perfect mathematical machines; responses vary between individual animals and tissues.
- Practice scientific data recording and analysis: Learning the rigor of maintaining lab notebooks, statistically analyzing data, and drawing objective conclusions.
In modern pharmacology laboratories, experiments may involve:
- Isolated tissue preparations: (Ex vivo) Testing drugs on organs kept alive outside the body.
- Animal experiments: (In vivo studies) Measuring parameters like blood pressure, behavior, or toxicology in a whole living animal.
- Computer simulation experiments: (In silico) Using advanced software to simulate biological responses without using living tissues.
- Drug bioassays: Determining the concentration or potency of a substance by measuring its biological response relative to a standard.
- Pharmacokinetic studies: Tracking drug absorption and elimination rates by taking serial blood or urine samples over time.
Introduction to Pharmacology Laboratory Instrumentation
Instrumentation is the lifeblood of experimental pharmacology. High-quality, properly calibrated instruments are absolutely essential for the accurate measurement, recording, and analysis of drug effects.
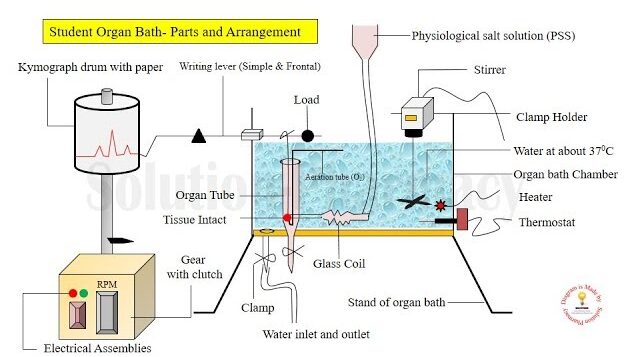
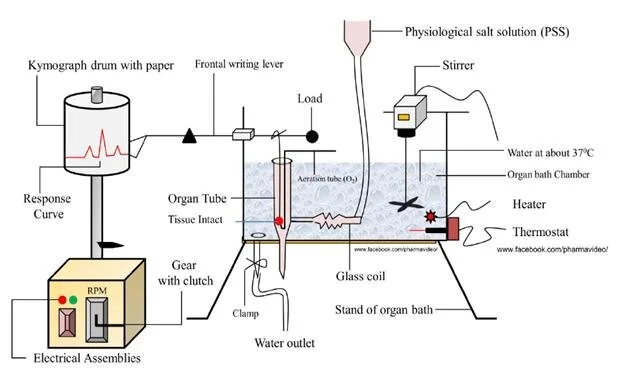
1. The Organ Bath System
The organ bath is a classic and foundational apparatus used to study the physiological effects of drugs on isolated tissues. By removing a tissue and placing it in a controlled environment, researchers can study local drug effects without interference from systemic reflexes or central nervous system control.
Typical tissues studied include:
- Ileum (part of the small intestine, commonly from a guinea pig or rat).
- Uterus (to study drugs that induce or inhibit labor contractions).
- Trachea (windpipe tissue to study bronchodilators used in asthma).
- Aorta (blood vessel tissue to study vasoconstriction and vasodilation).
- Heart muscle (atria or ventricles to study drugs affecting heart rate and contractility).
Components of a Student Organ Bath Assembly:
- Tissue Chamber (Organ Tube): A specialized inner glass tube where the isolated tissue is suspended. It contains a physiological salt solution (PSS) that mimics the body's natural fluids (e.g., Tyrode's or Krebs solution) to keep the tissue alive.
- Outer Water Bath: A larger chamber filled with water that surrounds the inner tissue chamber.
- Temperature Control (Thermostat & Heater): Maintains the water (and thereby the inner solution) at exact body temperature (~37°C for mammals). A stirrer ensures uniform temperature distribution.
- Aeration System (O2/CO2): Tissues require oxygen to survive. An aeration tube delivers gas (often "carbogen" - 95% Oxygen and 5% Carbon dioxide) directly into the physiological solution. The bubbling also helps mix the drug.
- Tissue Holder and Hooks: The bottom of the tissue is tied to a fixed hook (aeration tube base), while the top is tied via a fine thread to a transducer or writing lever.
- Transducer / Recording System: Detects the mechanical movement or tension of the tissue and converts it into a readable format.
The organ bath allows for precise measurement of muscle contraction, muscle relaxation, drug potency, and the generation of dose-response curves.
Example: Effect of Acetylcholine on Guinea Pig Ileum.
A piece of guinea pig intestine is suspended in the bath. When Acetylcholine (a neurotransmitter) is added via a micropipette into the physiological solution, it binds to muscarinic receptors on the smooth muscle of the ileum, causing a rapid, measurable contraction. By adding increasing doses, a student can plot a dose-response curve.
2. Physiological Recording Systems
These systems are responsible for capturing the physical biological response (like a muscle twitch) and recording it for analysis.
- a) Kymograph (Classical Instrument)
The kymograph is a historically significant, mechanical instrument. It essentially records tissue contraction on a rotating drum wrapped with smoked paper.
- Principle: The physical, mechanical movement from a contracting tissue pulls a thread connected to a magnifying lever (e.g., a simple or frontal writing lever). The tip of the lever lightly touches a rotating drum covered in a layer of black soot (smoked paper). As the tissue contracts, the lever moves up and scratches away the soot, leaving a white line tracing the contraction wave.
- Historical Use: While largely replaced by digital systems today, it was historically the backbone of isolated tissue studies, muscle contraction experiments, and early physiology research.
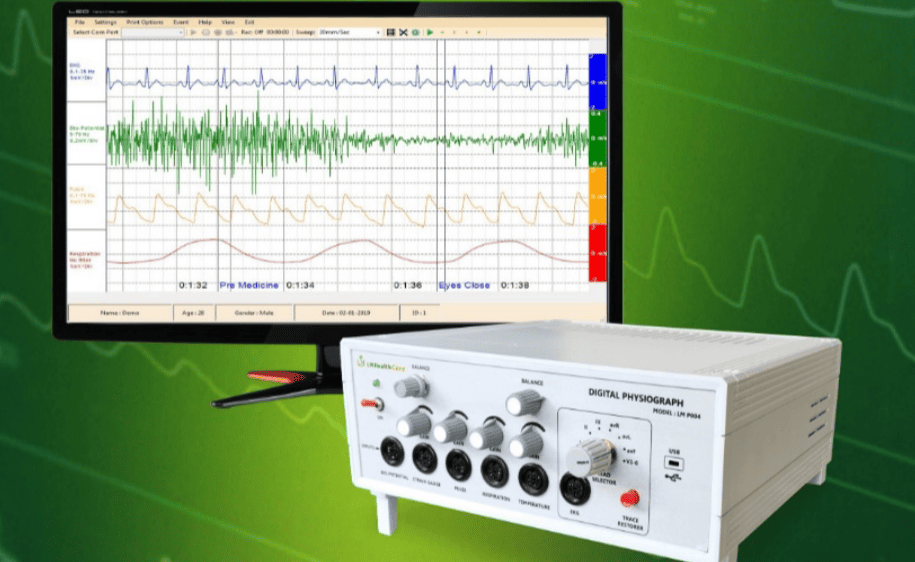
- b) Polygraph / Physiograph
These are the transitional electronic recording systems. Instead of a mechanical lever scratching paper, they use electronic sensors to record multiple physiological parameters simultaneously onto a scrolling chart paper or basic digital screen. They can concurrently record: Blood pressure, Heart rate, Muscle contraction, and Respiration depth/rate.
- c) Data Acquisition Systems (Modern Standard)
Modern laboratories have almost exclusively transitioned to highly sophisticated computer-based systems. Leading examples include systems manufactured by ADInstruments (PowerLab) and Harvard Apparatus.
- Components: Transducers (to capture the biological signal), Amplifiers (to boost the microscopic electrical signals), Data recording modules (hardware converting analog to digital), and Computer software (such as LabChart, which displays, stores, and analyzes data).
- Advantages: These modern systems allow for absolute real-time data recording, intricate digital analysis (calculating area under the curve, exact frequencies), and immediate graph generation for publication.
3. Transducers
A transducer is a critical intermediary device. Its primary function is to convert biological signals (mechanical force, pressure, displacement) into electrical signals that a computer or physiograph can understand and record.
There are two major types used in tissue baths:
- Definition: "Iso" = same, "metric" = length. These measure the force or tension generated by a muscle without allowing the muscle to change its length.
- Application: Used heavily in smooth muscle contraction studies and vascular tissue (blood vessel) experiments where the tension developed against a fixed resistance is the critical metric.
- Definition: "Iso" = same, "tonic" = tension. These measure the physical change in tissue length (shortening) during contraction while keeping the load/tension constant.
- Application: Used when studying the actual physical shortening of a tissue, such as a piece of gut pulling a lever upward.

4. Perfusion Pumps
Perfusion pumps are automated mechanical devices designed to ensure a steady, constant flow of physiological solutions or drugs to a tissue or animal over extended periods.
- Applications: Crucial in organ perfusion experiments (e.g., keeping an entire isolated heart continuously supplied with nutrients via the Langendorff setup) and continuous drug delivery studies.
Types include:
- Peristaltic pumps: Use rotating rollers to squeeze fluid through flexible tubing. Excellent because the fluid never touches the pump machinery, ensuring sterility.
- Syringe pumps: Slowly and mechanically depress the plunger of a loaded syringe to deliver highly precise, minute volumes of drugs (micro-infusions).
5. Analytical Instruments in Pharmacology Labs
Beyond tissue responses, modern pharmacology practicals frequently involve biochemical and analytical chemistry to determine drug concentration analysis within biological fluids.
- Spectrophotometers:
- Function: Used to highly accurately measure drug concentration by evaluating how much light a specific solution absorbs (based on the Beer-Lambert law).
- Example Type: UV-Visible Spectrophotometer (utilizes ultraviolet and visible light spectrums).
- Applications: Conducting drug assays, studying enzyme kinetics, and performing metabolic breakdown studies.
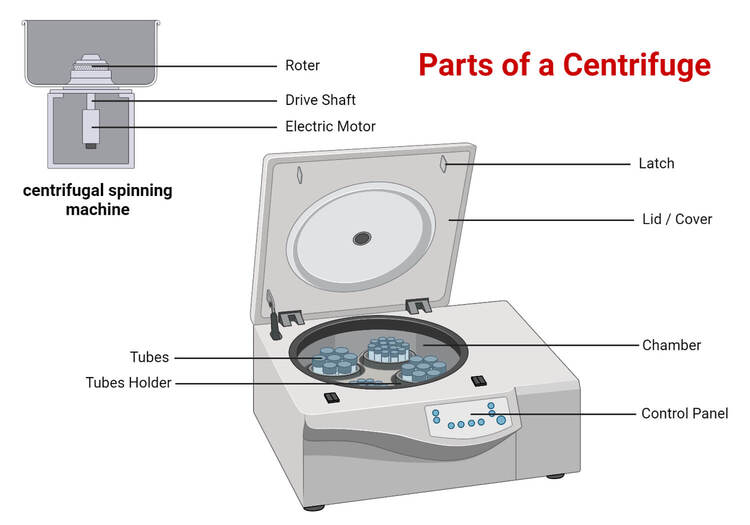
- Centrifuges:
- Function: Utilize rapid spinning (centrifugal force) to separate components of biological samples based on density.
- Applications: Separating clear blood plasma from heavy red blood cells, or preparing tissue homogenates (blended tissues) for molecular analysis.
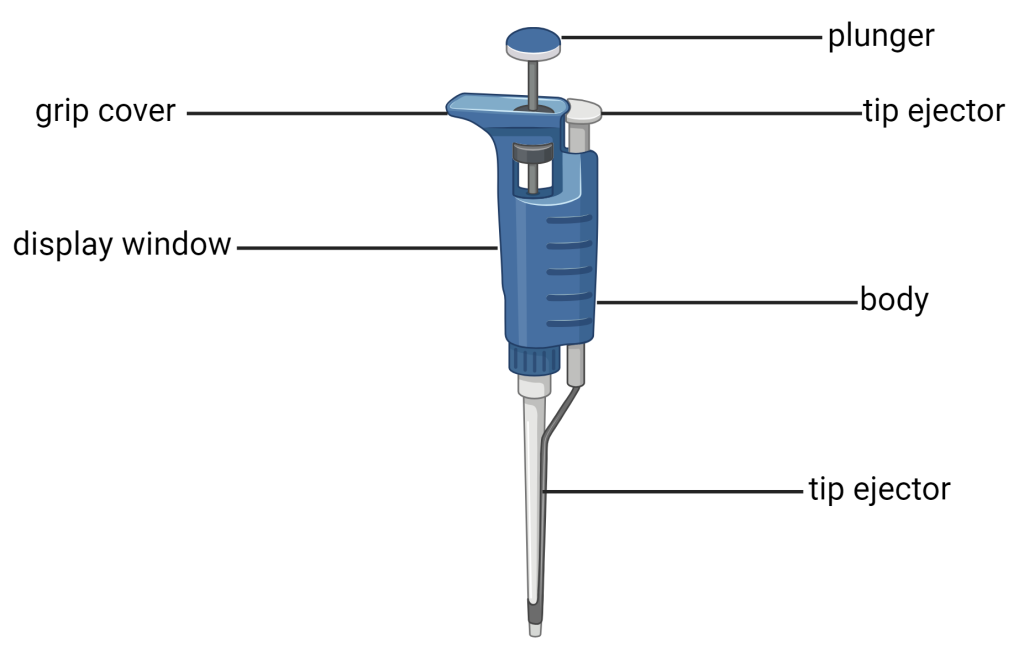
- Micropipettes:
- Function: Essential hand-held tools used for the extremely accurate measurement and transfer of very small liquid volumes, usually measured in microliters (µL). They are indispensable for adding exact drug doses to an organ bath.

5. Laboratory Safety and Ethical Considerations

Safety Equipment in Pharmacology Labs
Pharmacology labs deal with potent chemicals, biologically active drugs, and animal tissues. Safety is paramount to protect the researcher and the environment. Standard safety equipment includes:
- Fume hoods: Ventilated enclosures used to safely handle volatile toxic chemicals, preventing inhalation of hazardous vapors.
- Personal Protective Equipment (PPE): Specifically, nitrile gloves to prevent skin absorption of drugs, and heavy cotton lab coats to protect clothing and skin from spills. Safety goggles protect the eyes.
- Biohazard containers: Specially marked, puncture-proof bins (often red or yellow) for the safe disposal of biological tissues, blood-contaminated items, and sharp objects (needles/scalpels).
- Animal handling equipment: Specialized cages, thick gloves, and restraints to safely handle live animals without causing stress to the animal or injury to the handler.
- Emergency wash stations: Eye-wash basins and full-body safety showers to immediately dilute and flush away accidental chemical splashes.
These elements are strictly essential for the safe handling of drugs, hazardous chemicals, and biological samples.
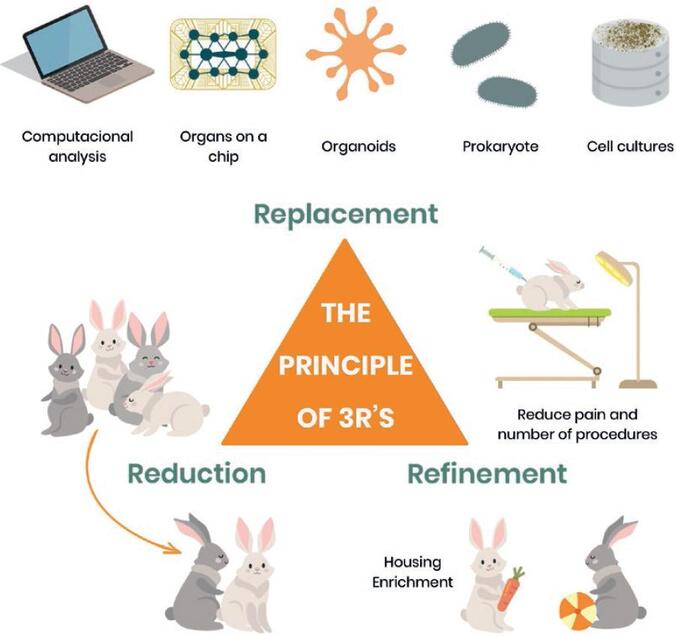
Ethical Considerations in Pharmacology Practicals
The use of live animals in science is a serious ethical issue. Modern pharmacology is strictly governed by ethical boards and humane principles. Any animal experiment must follow the internationally recognized framework known as The 3Rs Principle:
The 3Rs Principle
- Replacement: The absolute first step is to question if an animal is needed at all. Researchers must use alternative methods where possible, such as cell cultures (in vitro) or computer models.
- Reduction: If animals must be used, the experiment must be statistically designed to minimize the number of animals required to obtain valid, scientifically significant data.
- Refinement: Experimental procedures must be optimized to minimize animal suffering, pain, and distress. This includes proper housing, adequate anesthesia, and humane endpoints.
The Rise of Computer Simulations
In many modern educational institutions, to adhere to the principle of Replacement, computer simulations are increasingly used to entirely replace animal experiments for undergraduate teaching.
A prime example of this is ExPharm (and similar pharmacology simulation software). These programs allow students to administer "virtual drugs" to simulated tissues (like a virtual rat intestine or dog blood pressure model) on a screen. They generate realistic physiological graphs and data, allowing students to learn dose-response concepts and practical analysis without sacrificing a single animal life.
Introduction to Basic Pharmacology Read More »